276
Botanical Studies, Vol. 53, 2012
and C. tenuiflora throughout their geographic ranges in Taiwan are characterized. Two native sibling species, C. hengchunensis Chang and C. kissi from the section Paracamellia (Ming, 2000; Vijayan et al., 2009) which C. brevistyla and C. tenuiflora belong to, are also compared. Many previous studies show that plants usually exhibit a morphological plasticity in response to environmental
conditions (Thompson, 1991; Ro9as et al., 1997) and that frequently causes taxonomic problems. Since C. brevistyla and C. tenuiflora are widely distributed, a test of the correlation between morphological variation and geographical location of the two taxa is also conducted in this study.
MATERIALS AND METHODS
Materials
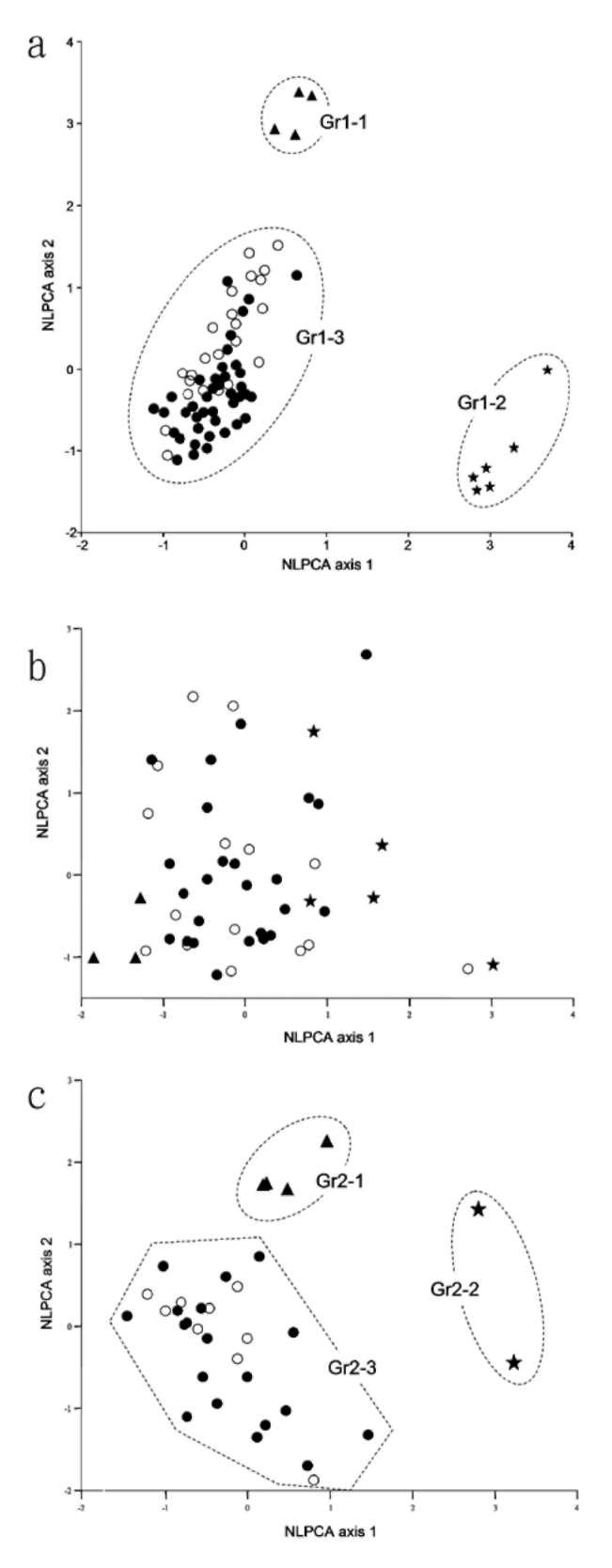
For scoring morphological characters, specimens deposited in the herbaria were examined (Appendix 1). Each collection was designated as an operational taxonomic unit (OTU). Duplicate specimens were treated as the same OTU. Scientific names given on the labels of specimens were tentatively used. A total of 149 specimens were examined, but those without sufficient measureable or label information were excluded from the data analyses. Finally, 74 specimens (OTUs), including 24 specimens of C. brevistyla, six of C. hengchunensis, four of C. kissi and 40 of C. tenuiflora, were included (Appendix 1). The specimens of C. brevistyla and C. tenuiflora represent their distribution range in Taiwan (Figure 1). Although type specimens of C. brevistyla and C. tenuiflora were not included in the study because they did not provide enough measurable traits, specimens from the type locality were used (e.g. C. brevistyla, M. H. Su 679 and 681, TAI; C. tenuiflora, P. F. Lu 7175, TNU; Appendix 1). The small sample sizes of C. hengchunensis and C. kissi were due to their limited distribution and small population sizes in Taiwan (Figure 1).
Characters analysis
Thirty-three characters were chosen on the basis of the variation mentioned in the literature and our own judgement: 12 leaf characters, 16 floral characters and five fruit characters (Tables 2-4). For each specimen, five mature and healthy-looking leaves were scored and averaged. The measurements of floral and fruit characters were averaged from one to three replicates, depending on the condition of the specimen.
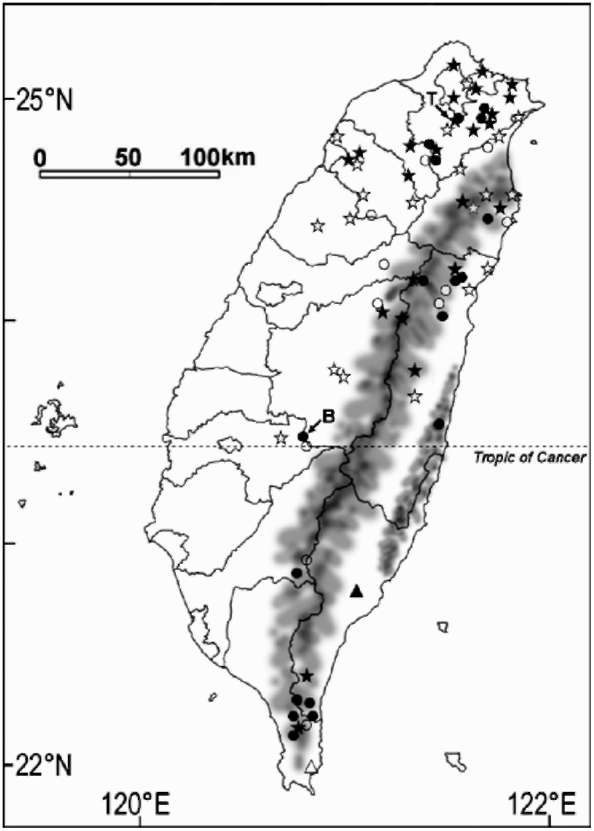
Figure 1. Distribution of the populations of Camellia brevistyla and C. tenuiflora in Taiwan. Solid circle, C. brevistyla sampled. Open circle, C. brevistyla not sampled. Solid star, C. tenuiflora sampled. Open star, C. tenuiflora not sampled. Solid triangle, C. kissi. Open triangle, C. hengchunensis. Arrow marked with B, collection site of type specimen of C. brevistyla. Arrow marked with T, collection site of type specimen of C. tenuiflora. Data were sourced from HAST, PPI, TAI, TAIF, TNM, and TNU.
|
Table 1. A comparison of the useful taxonomical features of Camellia brevistyla and C. tenuiflora cited in previous reports.
|
||||
Source
|
C. brevistyla
|
C. tenuiflora
|
||
Yamamoto and Mori, 1934 Secondary veins inconspicuous.
|
Secondary veins flat or stressed above, conspicuous beneath.
|
|||
|
Keng, 1950
|
Styles 4, 1 mm long, connate at base.
|
Styles 3, 3 mm long, free at upper portion or entirely connate at the base.
|
||
|
Liu and Lu, 1967
|
Secondary veins inconspicuous.
|
Secondary veins 5-8 pairs, conspicuous, slightly elevated on both surfaces.
|
||
|
Li, 1976
|
Styles 1-1.5 mm long, connate at base.
|
Styles 3 mm long, free at upper portion or entirely connate.
|
||
|
Ying, 1995
|
Leaf apex obtuse; styles 3-fid.
|
Leaf apex acute; styles shallowly 3-fid or entirely connate.
|
||
|
Hsieh et al., 1996
|
Styles 4, 1-1.5 mm long; leaves rhombic-oblong, midribs pubescent above.
|
Styles 3, more than 2.5 mm long; leaves obovate-elliptic, midribs almost glabrous above.
|
||