Bot. Bull. Acad. Sin. (1995) 36: 181-188
Hwang and Chen Anatomical changes in Kandelia candel
Anatomical responses in Kandelia candel (L.) Druce seedlings growing in the presence of different concentrations of NaCl
Yuan-Hsun Hwang1 and Shuh-Chun Chen
Institute of Botany, Academia Sinica, Taipei 115, Taiwan, Republic of China
(Received January 24, 1995; Accepted June 7, 1995)
Abstract. We studied the histology of leaves and roots of Kandelia candel seedlings growing in various salinities (0_550 mM NaCl). The structural changes found were related mainly to growth status. The plants grew best in the presence of 50 and 100 mM NaCl, with greater leaf area and thickness, higher ratios of palisade to leaf thickness, and higher stele to root cross-section-area proportions. Lignification and/or suberization of the cell wall appeared farther from the root apex. Structural modifications associated with adaptation to a saline environment were not found in this studyphysiological regulation may be more important.
Keywords: Anatomical changes; Kandelia candel; Mangrove; NaCl; Rhizophoraceae; Salinity.
Introduction
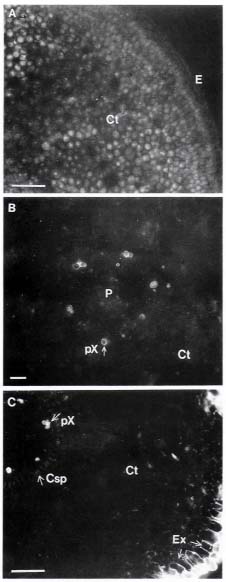
Plants that tolerate high salinity environments are termed halophytes. They always evolve structural modifications and physiological adaptations (Flowers et al., 1986). Some of the structural changes that occur in those halophytes that lack specialized salt-excretion organs such as salt glands or salt bladders include increased succulence, changed number and size of stomata, thickened cuticle, inhibited differentiation, and changed diameter and number of xylem vessels. There may also be extensive development of tyloses and earlier occurrence of Casparian strips and lignification (Flowers et al., 1986; Poljakoff-Mayber, 1975). Depending on the species, these changes may be either adaptations to salinity stress or signs of damage and disturbance of the normal life processes (Poljakoff-Mayber, 1975).
The mangrove Kandelia candel (L.) Druce grows in the intertidal zone along estuarian river banks, where the tidal salinity ranges between 10 and 36 parts per thousand (ppt) (Hwang, 1983). The optimal salinity for the growth of K. candel seedlings, however, is between 0 and 15 ppt (ca. 257 mM NaCl) (Hwang and Chen, 1995). The basic anatomical structures of this species have been studied by Chiang (1984). Structural changes were noticed when seedlings collected from saline environment in the field were grown in freshwater in a greenhouse for more than one year. These plants showed decreased thickness of leaves and cuticle layers, reduced tannin content in the tannic and palisade cell layers, increased intercellular spaces, and increased ratio of spongy cell layers to leaf (Chiang, 1984). The anatomical structure of K. candel changes in response to different salinity conditions, but whether these
changes are related to the development of salt tolerance was not clear. The present study investigates further the relationship between anatomical change and salt tolerance in K. candel.
Materials and Methods
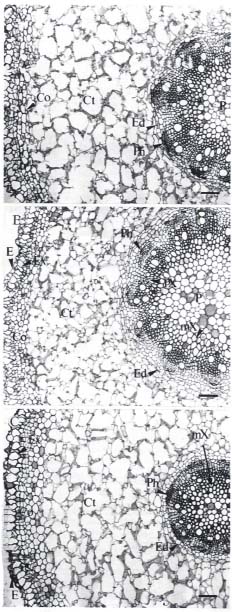
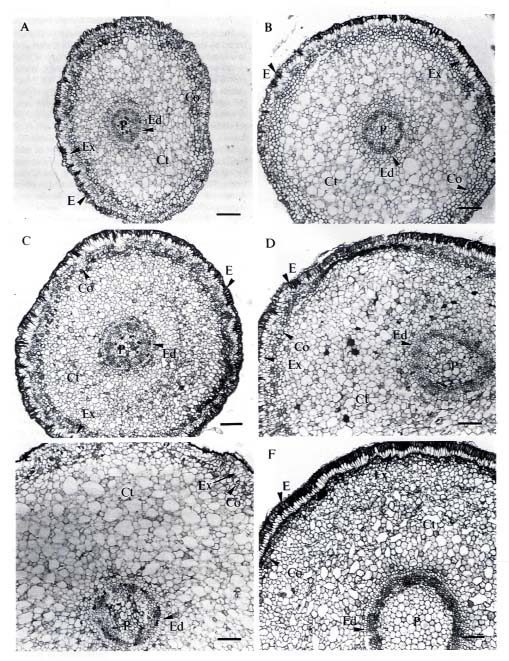
Mature propagules of Kandelia candel (L.) Druce, collected from mature trees growing along the Tamshui River, Taipei, Taiwan, (121° 26' E, 25° 9' N), were cultivated on sand in pots, partially submerged in nutrient solutions. The culture solution was a modified Hoagland's solution, with NaCl added at the beginning to form 0, 50, 100, 250, 400 and 550 mM NaCl solutions. Details of the culture method were described by Hwang and Chen (1995). We used mature leaves and roots of four-month-old plants for our study, except for the autofluorescence study, which used root tissues of one-month-old plants.
Light Microscopy
Leaf area was measured with a leaf-area meter (model Li-3000A, LI-COR Inc., Lincoln, Nebraska, USA). Leaf discs (7 mm diameter) were cut with a paper punch along the mid-rib at the central part of mature leaves. Segments (0.5 cm long) of mature root were cut 2 cm behind the hypocotyl, and segments (0.5 cm long) of root tip were cut 0.5 cm behind the apex. The segments were fixed for 8 h in FAA (37% formalin, acetic acid, 50% alcohol = 5:5:90). The materials were then dehydrated through a TBA (tertiary butyl alcohol) series and embedded in paraplast (Oxford Labware, Maryland, USA). Cross sections (10 µm) were cut with a rotary microtome (Leitz, West Germany) and stained with 1% safranin O and 0.5% fast-green (Johansen, 1940).
1Corresponding author.