Bot. Bull. Acad. Sin. (1995) 36: 215-221
Yang et al. Chlorophyll and light
Light-sensitivity of chlorophyll formation in the leaves of Ficus microcarpa cv. Golden-leaves
Chi-Ming Yang1,3, Jen-Chieh Hsu1, and Yung-Reui Chen2
1Institute of Botany, Academia Sinica, Taipei, Taiwan 115, Republic of China
2Department of Botany, National Taiwan University, Taipei, Taiwan 106, Republic of China
(Received March 4, 1995; Accepted July 17, 1995)
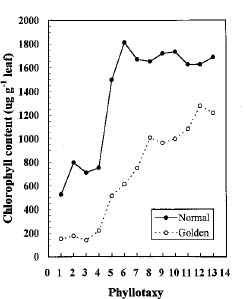
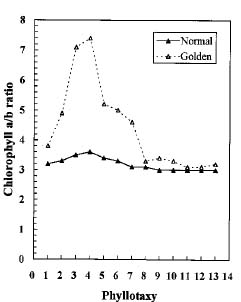
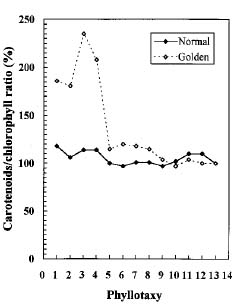
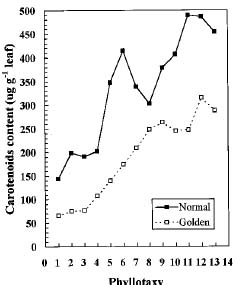
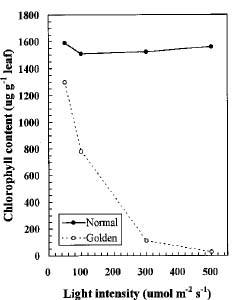
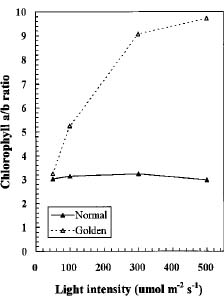
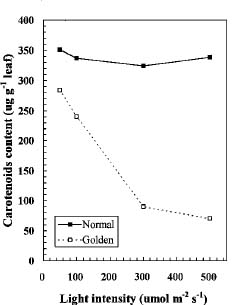
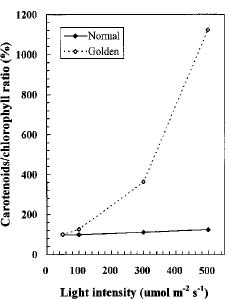
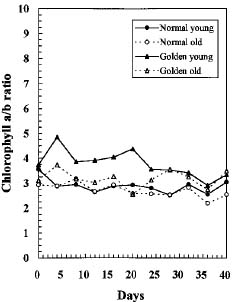
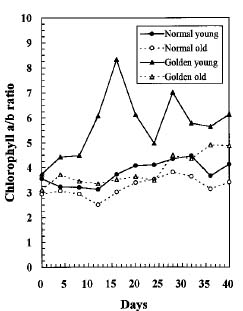
Abstract. We compared the influence of light intensity and illumination time on the chlorophyll (Chl) and carotenoid content, and on the Chl a/b and carotenoid/Chl ratios of leaves from the same shoot of normal and of Golden-leaves fig (Ficus microcarpa cv. Golden-leaves). Golden-leaves fig possesses light-sensitive leaves with various amounts of Chl and carotenoids and various ratios of Chl a/b and carotenoids/Chl under the same illumination. It appears that the deficiency of Chl is not directly associated with the loss of carotenoid, but the alteration of Chl a/b ratios might be related to that of the carotenoids/Chl ratio. It is likely that the relationship between synthesis of Chl b and carotenoids is more intimate than that of Chl a and carotenoids. The unique characteristics of Golden-leaves fig makes this plant a useful tool for studying the photobiochemistry and photomorphogensis of higher-plant chloroplasts.
Keywords: Carotenoid; Chlorophyll; Golden-leaves fig; Illumination duration; Illumination intensity.
Introduction
Chl-deficient mutants have been widely used to study the biosynthetic pathway of Chl and the biogenesis of the photosynthetic apparatus in higher plants (Somerville, 1986). These mutants have been reported in barley, maize, pea, sweetclover, wheat, Chlamydomonas reinhardtii (see references in Markwell et al., 1986), rice (Terao et al., 1985), soybean (Droppa et al., 1988), sugar beet (Abadia et al., 1985), and Arabidopsis thaliana (Hirono and Redei, 1963). They are generally divided into two groups: Chl b-lacking mutants, which contain no detectable Chl b, and Chl b-reduced mutants, which contain reduced amounts of this pigment (King, 1991). Some of these mutants are sensitive to light ( Hopkins et al., 1980; Markwell et al., 1986; Allen et al., 1988; Greene et al., 1988) and others are sensitive to temperature (Markwell et al., 1986). Among the temperature-sensitive mutants, fewer mutants were found to be sensitive to light intensity and/or photoperiod (Yang et al., 1993). Temperature sensitivity has been proposed to be a general phenomenon among the Chl-deficient mutants (Yang et al., 1990; Markwell and Osterman, 1992) that may attributable to blocked Mg-chelatase activity (Falbel and Staehelin, 1994). According to their response to the growth temperature, Chl-deficient mutants can be placed within two major biochemical phenotypes: 1) those mutants that adjust only their Chl content and 2) those that simultaneously adjust Chl content and Chl a/b ratio (Yang et al., 1993).
In a previous study, Golden-leaves fig was found to be sensitive to temperature (Chen and Yang, 1995) and to
possess light-sensitive leaves containing various amounts of Chl and having various Chl a/b ratios on the same shoot under the same illumination (Yang et al., unpublished data). In this report, we further study these traits and show that this plant is sensitive to intensity and duration of illumination.
Materials and Methods
Normal and Golden-leaves (Ficus microcarpa cv. Golden-leaves) fig plants about one-year old and 50-cm tall were purchased from a local nursery. Plants were grown in sterile soils in a controlled-environment growth chamber with a 12/12 h photoperiod, 25°C temperature, 60_70% relative humidity, and various illumination intensities.
Following the extraction of liquid-nitrogen-frozen leaves with 80% (v/v) acetone, Chl concentration and Chl a/b ratios were determined according to the method of Porra et al. (1989). The concentration of total carotenoids was measured by the method of Kirk and Allen (1965). Room temperature absorbance was obtained with a Hitachi U2000 UV-visible spectrophotometer.
Three determinations were made for each experiment and similar patterns were obtained. Only one of the patterns is presented in this paper.
Results and Discussion
The various developmental stages of the leaves of Golden-leaves fig can be classified by their color (Chen and Yang, 1995). The new and yellow leaves on the top are in the young stage (the first four or five leaves), yel
3Corresponding author.