Bot. Bull. Acad. Sin. (1996) 37: 31-34
Yang and Chen Grana stacking in chloroplast
Grana stacking is normal in a chlorophyll-deficient LT8 mutant of rice
Chi-Ming Yang1,3 and Hsiu-Yu Chen2
1Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan 115, Republic of China
2Department of Agronomy, National Chung-Hsing University, Taichung, Taiwan 402, Republic of China
(Received May 12, 1995; Accepted August 14, 1995)
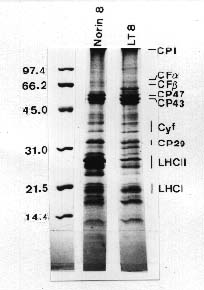
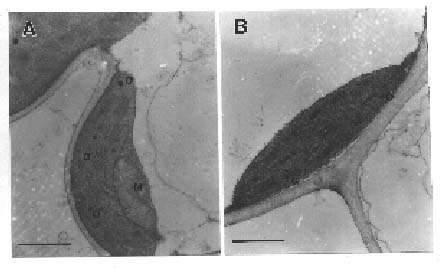
Abstract. Thylakoid proteins and chloroplasts isolated from chlorophyll-deficient rice mutant LT8 and its Norin 8 parent type (Oryza sativa) were compared using SDS-PAGE and electron microscopy. The LT8 mutant contained approximately 37% of the amount of chlorophyll found in the Norin 8; the ratio of chlorophyll a/b for LT8 and Norin 8 were 6.4 and 3.2, respectively. SDS-PAGE profiles showed that LT8 mutant contained approximately 10% of the amount of LHCII polypeptide found in the Norin 8, although both biotypes produced similar levels of LHCI and other thylakoid proteins. Electron microscopy showed that the grana stacks were similar in the LT8 mutant and Norin 8, suggesting that LHCII is not the only factor regulating the grana stacking of thylakoid membrane.
Keywords: Chlorophyll-deficient mutant; Grana stacking; LHCII; Oryza sativa.
Abbreviations: Chl, chlorophyll; CF, coupling factor; Cyf, cytochrome f; PSI, photosystem I; PSII, photosystem II; LHCI and LHCII, light-harvesting complexes associated with PSI and PSII, respectively; SDS-PAGE, sodium dodecyl sulfate polyacrylamide gel electrophoresis.
Introduction
Grana stacks are predominant ultrastructural characteristics of thylakoid membranes in higher plants and green algae. It has been postulated that grana stacking is mediated by the surface charge density of thylakoid membrane (Barber, 1982), which is regulated by the phosphorylation and dephosphorylation of LHCII apoproteins in the thylakoid membrane (Staehelin and Arntzen, 1983; Murphy, 1986; Bennett, 1991; Allen, 1992ab). If this is the case, the chloroplast mutants that are Chl b-deficient and contain little or no LHCII complex should exhibit poorly-developed or no grana stacks in the thylakoidas in etiolated chloroplasts. A Chl b-lacking ch5 mutant (U374) of sweetclover, however, exhibits normal grana to an extent similar to its wild type, indicating that components other than LHCII are involved in grana stacking (Nakatani and Baliga, 1985). It has been suggested that the intraplastidial lamellar system in the plastids of a Chl b-lacking mutant in barley reorganized to form macrograna and optimize energy distribution between PSI and PSII (Ouijja et al., 1988).
In this report, we show that LT8 mutant, although containing little LHCII apoprotein, developed a normal grana stacking in the thylakoid membranes.
Materials and Methods
Seeds of rice mutant LT8 and its Norin 8 parental type were germinated and grown in a greenhouse for 5 weeks
in a soil-vermiculite mixture under natural light. Thylakoid membranes were obtained as previously described (Markwell, 1986). The concentrations of Chl and protein were determined using the method of Porra et al. (1989) and the Bio-Rad protein assay (Bradford, 1976), respectively. SDS-PAGE fractionation was performed with 12% acrylamide, using the method of Laemmli (1970).
For transmission electron microscopy, the central part of all leaves of both biotypes was collected and cut into small cubes (approx. 1 mm) in fixation buffer containing 2.5% glutaraldehyde. After incubation at 4°C for 2 h in 0.1 M cacodylate buffer (pH 7.0) containing 2.5% glutaraldehyde, the leaf samples were washed three times in plain buffer, postfixed in 1% osmium tetraoxide for 2 h, dehydrated through an ethanol series, infiltrated and embedded in Spurr's resin (Spurr, 1969), and then polymerized at 70°C for 8 h. Gold sections were collected by ultramicrotomy and stained with ethanol uranyl acetate and lead citrate. The thylakoid morphology was examined with a Hitachi H-600 transmission electron microscope at 75 kV.
Results and Discussion
SDS-PAGE Profiles
In normal green plants and green algae, about 50% of all Chl molecules are distributed in LHCII, which has a lower Chl a/b ratio than does LHCI (Murphy, 1986). Five-week-old LT8 mutant contained approximately 37% of the amount of Chl found in its parent type, Norin 8. Chl a/b ratios for LT8 and Norin 8 were 6.4 and 3.2, respectively
3Corresponding author.