Bot. Bull. Acad. Sin. (1996) 37: 61-87
Wang Systematics of Taiwanese Arisaema (Araceae)
The Systematic Study of Taiwanese Arisaema (Araceae)
Jenn-Che Wang
Department of Biology, National Taiwan Normal University,Wenshan, Taipei, Taiwan 11718, Republic of China
(Received August 11, 1995; Accepted November 14, 1995)
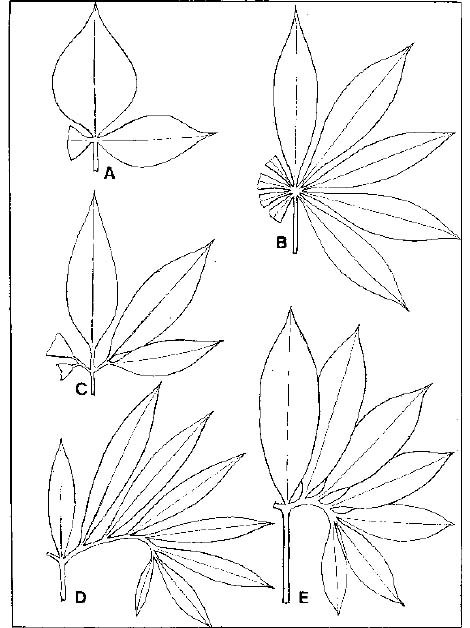
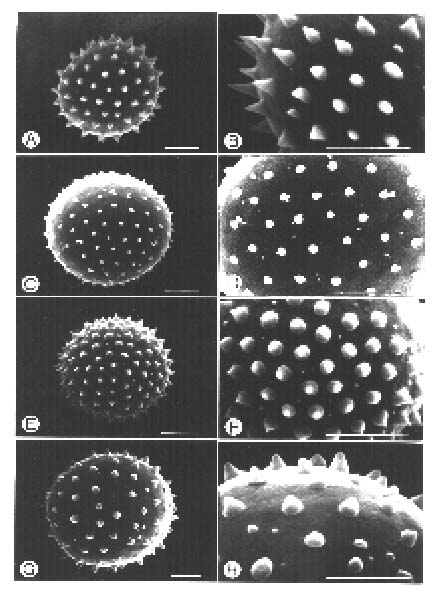
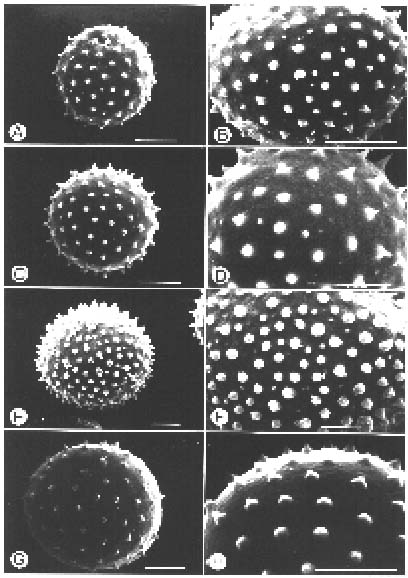
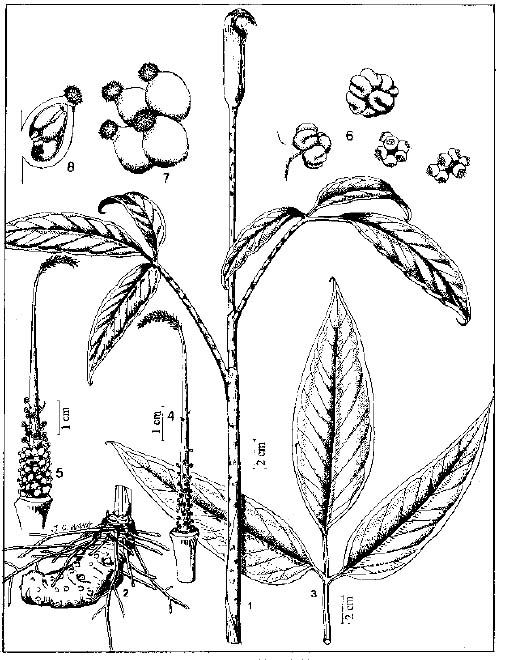
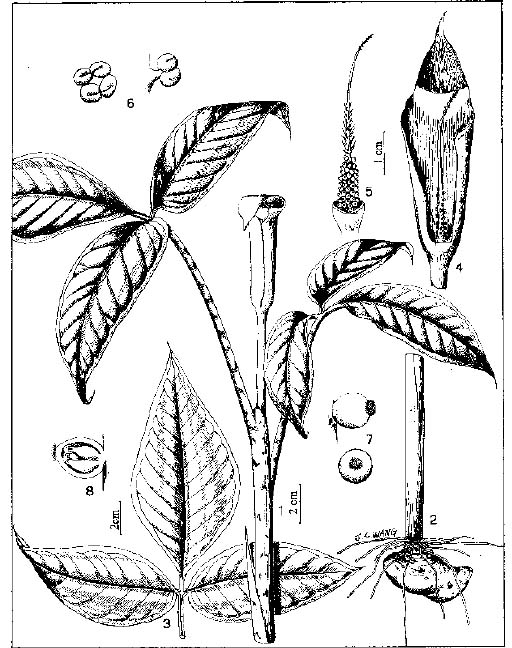
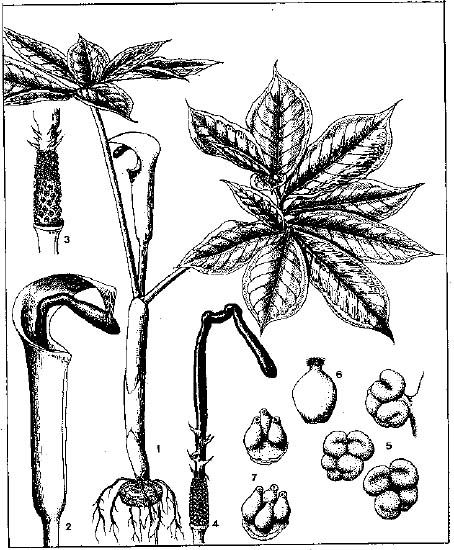
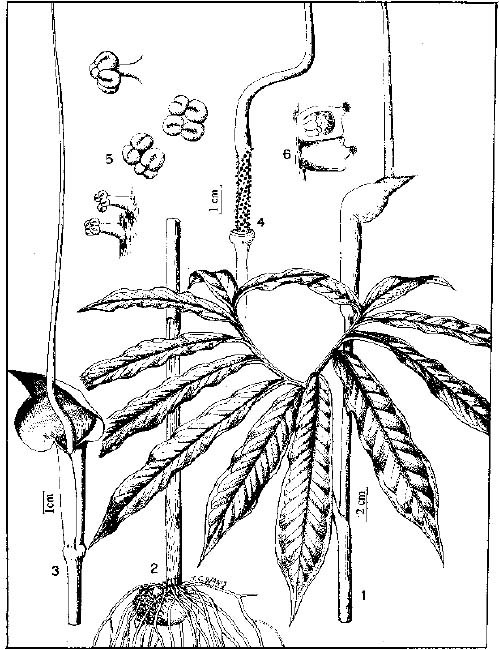
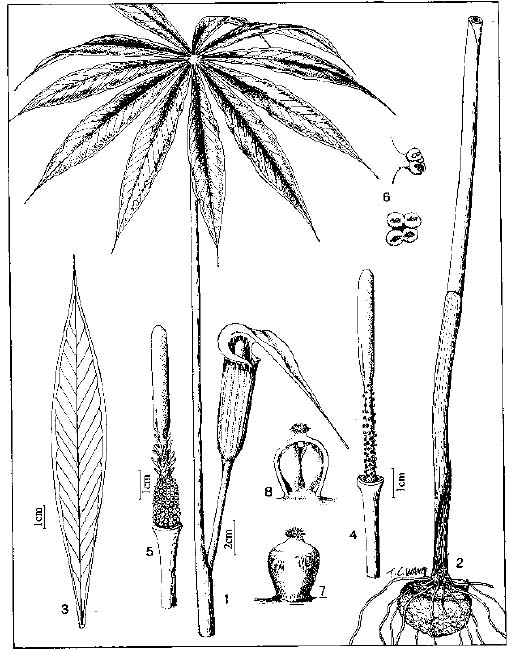
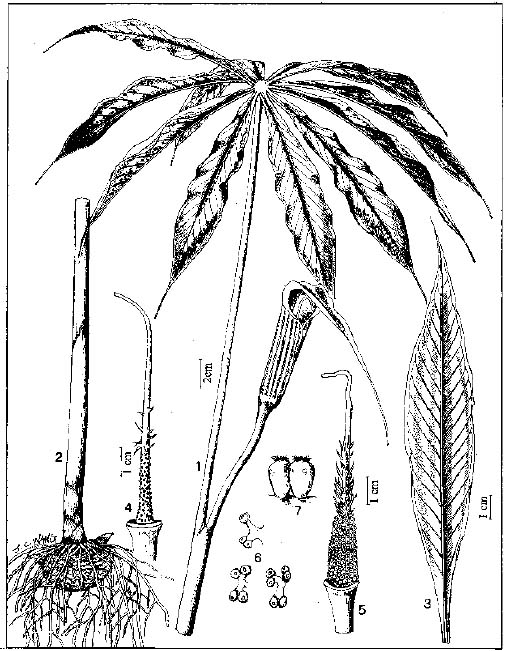
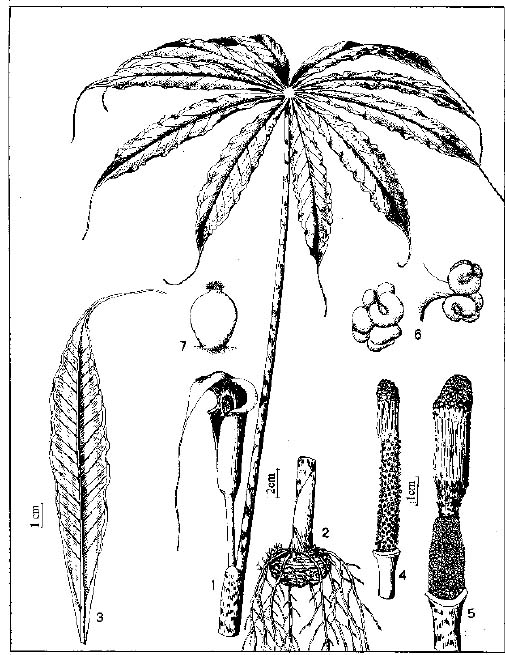
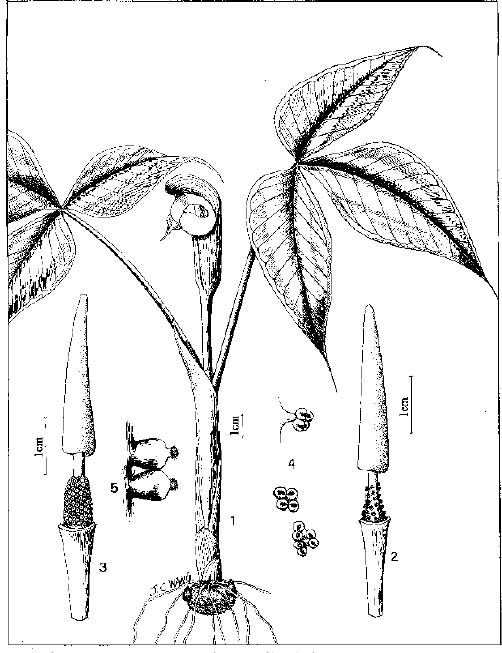
Abstract. The genus Arisaema (Araceae) in Taiwan is revised based on comparative morphological, palynological, and cytotaxonomical studies. Nine species and one variety, including two new taxa (A. ilanense J. C. Wang sp. nov. and A. thunbergii ssp. autumnale J. C. Wang, J. Murata & Ohashi ssp. nov.), are recognized. By the re-collection of fresh materials and a detailed comparison, A. matsudai Hayata, which was previously reduced to synonymy under A. grapsospadix Hayata, is now treated as distinct. Rhizome shape, arrangement of leaflets, ratio of pseudostem to petiole length, and spadix-appendage morphology are all important characters in separating the Taiwanese taxa. Pollen morphology also displays interspecific variation in size, spinule length, and spinule density. Cytologically, all examined species have 28 chromosomes (diploid), but three of them are also found to have 56 chromosomes (tetraploid). The chromosome numbers 2n=28 of A. ilanense and A. thunbergii ssp. autumnale, and 2n=56 of A. formosanum and A. heterophyllum are determined for the first time. A key to the taxa along with descriptions, illustrations, distribution maps, and taxonomic notes are provided.
Keywords: Araceae; Arisaema; Chromosome number; Pollen; Systematics; Taiwan.
Introduction
The genus Arisaema consists of about 150 species (Murata, 1990a) with the center of distribution and differentiation in warm to cool temperate regions of Asia (Li, 1980; Murata, 1984). The genus was first established by Martius in 1831 based on the Himalayan species (Murata, 1990a). The most important early monograph was by Engler (1920), who divided the genus into 15 sections based predominantly on the floral morphology. His system was later revised by Hara (1971), who emphasized the arrangement of leaflets rather than floral morphology and rearranged the Himalayan species into 13 sections. Hara's system was adopted by Li (1979) in her study of the Chinese species. Recently, Murata (1984) re-examined the characters which were used in the previous studies, and with special reference to fresh materials, he found many new taxonomic characters. Based on extensive studies including gross morphology, chromosome numbers, and pollen morphology he proposed a revised system (Murata, 1984, emend 1991) in which the genus was divided into 11 sections.
The Taiwanese Arisaema have been studied by few taxonomists. Most species hitherto known were described by Hayata (1915, 1916, 1920), Hosokawa (1936), and Kitamura (1941). The first revision of the Taiwanese species was by Huang (1960), who recognized five species and two forms. His treatment was adopted with little modification by Liu & Huang (1978) in the Flora of Taiwan, first edition. Subsequently, four taxa, viz. A. formosanum var. bicolorifolium Huang, A. taiwanense J. Murata, A. taiwanense var. brevipedunculatum J. Murata and A. thunbergii ssp. urashima (Hara) Ohashi & J. Murata, have been reported from Taiwan (Huang, 1982; Murata, 1985; Wang, 1992).
In the present paper an intensive revision is made based on comparative morphological, palynological, and cytological studies. As a result, nine species and one variety, including two new taxa, are recognized in Taiwan. All of these taxa are classified into five sections based on the system proposed by Murata (1984, 1991).
Materials and Methods
Materials used in the present study were collected from the field throughout Taiwan. Most material was pressed and dried for voucher specimens deposited in the Herbarium, Department of Biology, National Taiwan Normal University (TNU). Voucher specimens for pollen observation and chromosome counts were also deposited in TNU. Living material for the study was cultivated in the shade house of the Department of Biology, National Taiwan Normal University. In addition, specimens preserved in the following herbaria were examined: HAST: Herbarium, Institute of Botany, Academia Sinica, Taipei; PPI: Herbarium, Department of Forest Resource Management and Technology, National Pingtung Polytechnic Institute; TAI: Herbarium, Department of Botany, National Taiwan University; TAIF: Herbarium, Taiwan Forestry Research Institute; TNU: Herbarium, Department of Biology, National Taiwan Normal University.
Although most type specimens were unavailable, photographs of type specimens preserved in the Herbarium of Tokyo University (TI) (Ohashi, 1981) were examined by the author.
Pollen grains for scanning electron microscopic (SEM) study were collected from fresh anthers and prepared by the method proposed by Erdtman (1952). The acetolyzed grains were dehydrated through ethanol series and