Bot. Bull. Acad. Sin. (1996) 37: 107-111
Chen and Pan Assay of superoxide dismutase activity
Assay of superoxide dismutase activity by combining
electrophoresis and densitometry
Ching-Nen Chen and Shu-Mei Pan1
Department of Botany, National Taiwan University, Taipei, Taiwan 10764, Republic of China
(Received November 20, 1995; Accepted March 2, 1996)
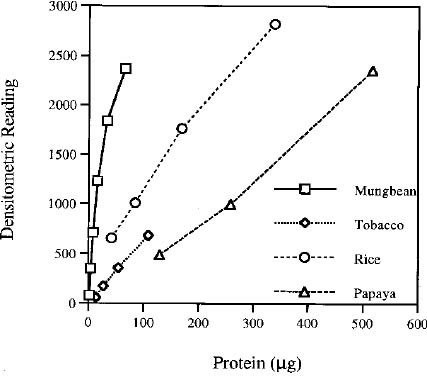
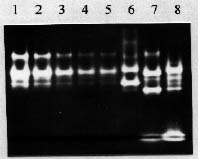
Abstract. A modified technique was developed to assay superoxide dismutase (SOD) activity by combining polyacrylamide gel electrophoresis and densitometry. After electrophoresis on native polyacrylamide gels, the negative banding corresponding to the SOD activity was visualized by soaking the gels in nitroblue tetrazolium then riboflavin, and finally exposing to light. Effects of the banding of SOD activity induced by different soaking durations and light intensities were evaluated in this system. The optimal soaking duration was determined to be 15 min for each of the two soaking steps, while the optimal exposure was 30 mEm-2s-1 for 15 min. The gels were then immediately scanned with a laser densitometer, and the readings of the samples corresponding to their total SOD activity were obtained by processing the image. A standard curve was prepared with a serial dilution of partially purified SOD, whose activity was previously determined by using a spectrophotometric method. The total SOD activity of an unknown sample could be obtained by interpolating its reading to the standard curve. The activity of a single SOD isozyme of a sample could also be obtained with the same procedure. The technique was ten times more efficient than the spectrophotometric method. The interference coming from non-SOD substances in the crude extract could be removed by electrophoresis. The standard deviations of the SOD activity of the crude extracts from rice seedlings, papaya, and tobacco leaves measured with the technique were less than 9%, 7%, and 8% (for each n = 6, on 6 gels), respectively.
Keywords: Densitometry; SOD isozyme; Superoxide dismutase.
Abbreviations: DETAPAC, diethylenetriamine pentaacetic acid; ED, electrophoretic-densitometry; NBT, nitroblue tetrazolium; SOD, superoxide dismutase; TEMED, tetramethylenediamine.
Introduction
Superoxide radical (O2_) is generated as a by-product in aerobic organisms from a number of physiological reactions such as the electron flow in the chloroplasts and mitochondria and from some redox reactions in cells. It can react with hydrogen peroxide (H2O2) to produce hydroxyl radical (OH_), one of the most reactive molecules in the living cells. Hydroxyl radical can cause the peroxidation of membrane lipids, breakage of DNA strands, and inactivation of enzymes in cells (for reviews, see Bowler et al., 1992; Mehdy, 1994). To ameliorate the damage caused by hydroxyl radical formed from superoxide radical and hydrogen peroxide, organisms have evolved mechanisms to control the concentration of the two reactants. Superoxide dismutase (SOD, EC 1.15.1.1) is a group of isozymes functioning as superoxide radical scavenger in the living organisms. The reaction of SOD is as follows:
2H+ + 2 O2_ ® H2O2 + O2
the produced hydrogen peroxide is then detoxified by catalase or peroxidase.
The expression of SOD genes is regulated both spatially and developmentally at least in maize (Zhu and Scandalios, 1993) and rice (unpublished data). SOD activity is also induced by diverse stresses (Bowler et al., 1992), presumably because of the increase in the concentration of superoxide radical in cells under those conditions. Obviously, SOD is an important enzyme family in living cells for maintaining normal physiological conditions and coping with stress. However, the study of SOD gene expression regulation at the end product level has been handicapped by the lack of a convenient method for quantifying the activity assay of the isozymes. Most of the work concerning SOD gene expression regulation has been based on RNA gel blot analysis. Little is known about the activity change of the SOD isozymes of plants in the developmental course and in response to stresses. SOD activity is commonly assayed spectrophotometrically, e.g., the method first defined by McCord and Fridovich (1969) and modified by Oberley and Spitz (1985). But it is both labor-intensive and time-consuming. We have developed a convenient technique for assaying the SOD activity by combining electrophoresis and densitometry and have called it the ED scheme. This technique is based on Beauchamp and Fridovich's method (1971), but is quantitative rather than qualitative. The ED scheme is more than ten times more efficient than the spectrophotometric
1Corresponding author. Fax: 886-2-3918940; E-mail: pan@ccms.ntu.edu.tw.