Bot. Bull. Acad. Sin. (1996) 37: 113-119
Liu et al. Effect of light on indole-3-acetic acid in soybean hypocotyls
Effect of light on endogenous indole-3-acetic acid, peroxidase and indole-3-acetic acid oxidase in soybean hypocotyls
Zin-Huang Liu1, Ho-Yih Liu and Hwei-Yi Wang
Department of Biology, Institute of Life Sciences, National Sun Yat-Sen University, Kaohsiung, Taiwan 804, Republic of China
(Received August 8, 1995; Accepted January 8, 1996)
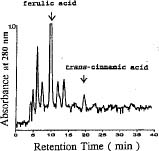
Abstract. Two days of light irradiance reduced the levels of endogenous indole-3-acetic acid (IAA) in soybean (Glycine max) hypocotyls. The key enzymes for IAA metabolic degradation are peroxidases and IAA oxidases. Peroxidase activity was enhanced about twofold by light. IAA oxidase activity also increased after irradiance. The lignin content and total phenolic compounds in soybean hypocotyls were enhanced 2_2.5 fold by light treatment. Light-induced inhibition of soybean hypocotyl growth might be due to the decline of endogenous IAA, which was degraded by IAA oxidases or peroxidases. The high levels of lignin were correlated with the increased peroxidase activity in light-treated tissues. Two phenolic compounds, ferulic acid and trans-cinnamic acid, which are precursors for lignin synthesis, greatly increased in light-treated soybean hypocotyls. This suggested that lignin synthesis catalyzed by peroxidase was enhanced during irradiance.
Keywords: Glycine max L.; IAA; Indole-3-acetic acid; Peroxidase.
Introduction
It is well known that light can inhibit the elongation of etiolated plant seedlings (Fletcher and Zalik, 1965; Jose and Vince-Prue, 1977; Ingram et al., 1983). Indole-3-acetic acid (IAA), a plant hormone, plays an important role in the promotion of hypocotyl elongation. Considerable attention has been paid to the possibility that light causes growth-inhibition by reducing the availability or effectiveness of IAA in the irradiated tissues (Iino, 1982; Jones et al., 1989; Jones et al., 1991). Free IAA can be modulated via several pathways, including IAA metabolism, synthesis and breakdown of conjugated IAA, and transport of IAA. One well known IAA metabolic pathway consists of oxidative decarboxylation of the side chain of IAA by IAA oxidase leading to the formation of either indole-3-methanol or 3-methylene oxindole (Grambow and Langenbeck-Sckwich, 1983; Beffa et al., 1990). Little is known concerning the correlation between levels of IAA and IAA oxidase activity during the processes of light-induced growth inhibition. One aim of this investigation was to examine the effect of light irradiance on soybean hypocotyl elongation through regulating the endogenous IAA levels which are modulated by IAA oxidase.
As the young seedlings in this study were exposed to light, their stems became more rigid. Hence the process of lignification in the cell wall may have occured. Numerous reports have indicated that particular peroxidases have the ability to catalyze polymerization of some aromatic alcohols to synthesized lignin (Bruce and West, 1989; Polle et al., 1994). In addition to examining the effect of light
on IAA levels, the effect of irradiance on lignification in soybean hypocotyl is also investigated.
Materials and Methods
Plant Materials
Soybean seeds (Glycine max) were obtained from the Asia Vegetable Research and Development Center, Taiwan. Dark-grown soybean seedlings were grown in vermiculite at 27 ± 0.5 °C for five days. Light-treated seedlings were grown in darkness for three days and then transferred to continuous light (16.7 W m-2) for two days. The apical region of each seedling's hypocotyl (2 cm) was excised after five days of growth for all experiments.
The Estimation of Indole-3-Acetic Acid
Extraction and partial purificationFive grams fresh weight of soybean hypocotyl segments were excised and immediately homogenized in 100 ml extracting solution with chilled motar and pestle (Guinnet et al., 1986; Dunlap and Guinn, 1989). The extracting solution containing 0.5 mg ml-1 butylated hydroxytoluene (BHT) in 80 ml of methanol and 0.5 mg ml-1 sodium ascorbate in 20 ml of distilled water prevented oxidation of IAA during extraction. The homogenates were filtered with suction through Whatman No.1 paper. Methanol was removed from the filtrate by rotary flash evaporation (RFE) at 35°C. The aqueous residue was adjusted to pH 8.0 with 0.2 N KOH and partitioned twice with an equal volume of ethyl acetate to remove phenolic compounds and other impurities. The ethyl acetate fraction was discarded and the residual ethyl acetate in aqueous phase was removed by RFE. The aque
1Corresponding author. Fax: 886-7-5319333.