Bot. Bull. Acad. Sin. (1996) 37: 127-131
Wei et al. Structure of extracellular polysaccharide
Structural studies of extracellular polysaccharide produced by Rhizobium fredii Tu6, a polysaccharide with nonasaccharide repeating units
Guor-Jien Wei1, Shuh-Chyung Song2, Liang-Ping Lin2, Guor-Rong Her1,3
1Department of Chemistry, 2Department of Agricultural Chemistry, National Taiwan University, Taipei, Taiwan, Republic of China
(Received October 7, 1995; Accepted February 14, 1996)
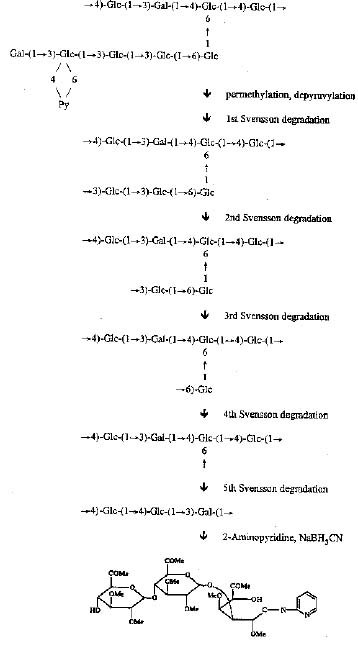
Abstract. Polysaccharide secreted by Rhizobium fredii Tu6 was investigated with composition analysis, linkage analysis, Svensson degradations, 1H-NMR, and desorption chemical ionization/mass spectrometry. The polysaccharide is composed of a nonasaccharide repeating unit (seven glucose, two galactose) with five sugar residues in the side chain and four sugar residues in the back bone. The terminal residue of the side chain is a galactose residue. The second residue from the terminal end is a 3-linked glucose substituted by a pyruvate group at the 4, 6-positions. Monosaccharide sequence was determined by Svensson degradations followed by linkage analysis. After removal of a pyruvate group from the methylated polysaccharide, the sugar residues in the side chain were removed sequentially by Svensson degradations. The reaction product of each degradation was then analyzed by methylation analysis. The polysaccharide secreted at pH 5 medium has a structure very similar to the polysaccharide secreted in the medium of pH 7 except that about 30% of the terminal galactose residues are also substituted at the 4, 6-positions by a second pyruvate group.
Keywords: Desorption chemical ionization/mass spectrometry (DCI/MS); Extracellular polysaccharide; Rhizobium fredii Tu6; Svensson degradation.
Introduction
Rhizobium species can infect leguminous plants and form nodules. The exopolysaccharides (EPS) excreted by the strains of Rhizobium play an important role in the infection of legumes and the Rhizobium / legume symbiosis (Phillip-Hollingsworth et al., 1989). Rhizobium fredii Tu6, a fast growing bacterium under neutral (pH 7) and acidic (pH 5) conditions, was isolated at Ping-Tong, Taiwan (Cheng and Lin, 1984). We have found a difference in morphological exhibition between neutral and acidic culture environments (unpublished data). We assume that EPS plays not only an important role in infection processes, but also a protective role in environmental adaptation. We now describe an investigation of the structure of the polysaccharide excreted by this bacterium. It is hoped that the investigation of the structure of EPS can provide some clues to the mechanism of infection and/or protection.
Materials and Methods
Isolation of the Polysaccharide
One and half day old (for pH 7) and two day old (for pH 5) cultures of R. fredii Tu6 were grown in B III medium (Bishop et al., 1976) and centrifuged. The polysac
charide was precipitated with cetyltrimethylammonium bromide. The precipitate was washed with ethanol, dialyzed against water, and lyophilized. The resulting crude exopolysaccharide was purified by a DEAE-sepharose column. Fractions containing exopolysaccharide were identified with a phenol-sulfuric acid test.
Proton NMR Spectroscopy
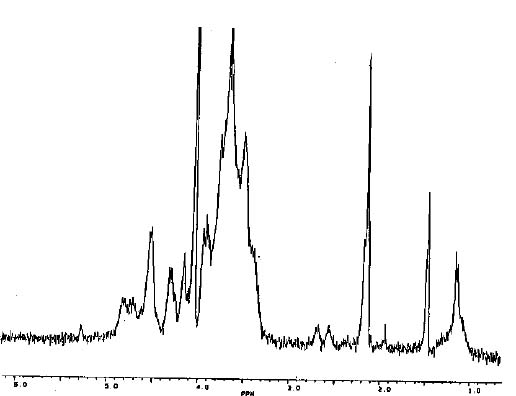
Samples for n.m.r. spectroscopy were lyophilized several times with D2O, and dissolved in D2O at a concentration of 1_3 mg/ml. Spectra were obtained with a Bruker 300-MHz instrument, with the probe heated to 60°C. The signal at d 1.47 was assigned to the methyl protons of pyruvic acid residues; the signal at d 2.13 was assigned to the methyl protons of O-acetyl groups; and the signals at d 2.57 and d 2.68 were assigned to the methylene protons of O-succinyl groups (Figure 1). The signal at d 1.17 remains to be assigned.
Desorption Chemical Ionization Mass Spectrometry
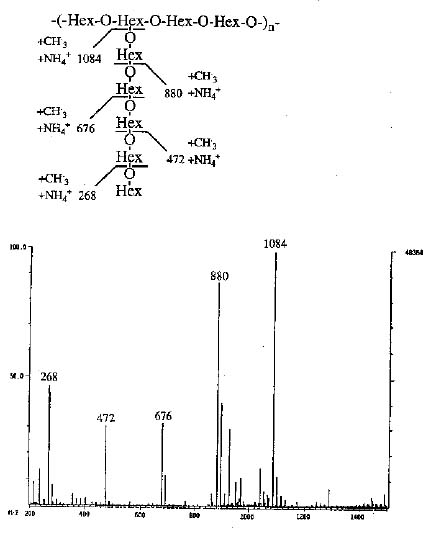
DCI/MS experiments were performed on a Finnigan TSQ-46C triplequadrupole mass spectrometer (Finnigan MAT, CA). Ammonia (from San-Fu Co., Hsinchu, Taiwan) was used as the reagent gas. The sample was air-dried on a platinum or rhenium emitter; the emitter was heated by a separated power supply at a heating rate of 50 mA/sec until the maximum current of 1.3 A was reached.
3Corresponding author. Fax: (02) 3638058.