Bot. Bull. Acad. Sin. (1996) 37: 133-140
Shiah et al. Light effect on phytoplankton photosynthetic
Light effects on phytoplankton photosynthetic performance in the southern East China Sea north of Taiwan
Fuh-Kwo Shiah1,2,4, Gwo-Ching Gong3, and Kon-Kee Liu1,2
1Global Change Research Center, National Taiwan University, Taipei, Taiwan, Republic of China
2Institute of Oceanography, National Taiwan University, Taipei, Taiwan, Republic of China
3Department of Oceanography, National Taiwan Ocean University, Keelung, Taiwan, Republic of China
(Received October 28, 1995; Accepted February 28, 1996)
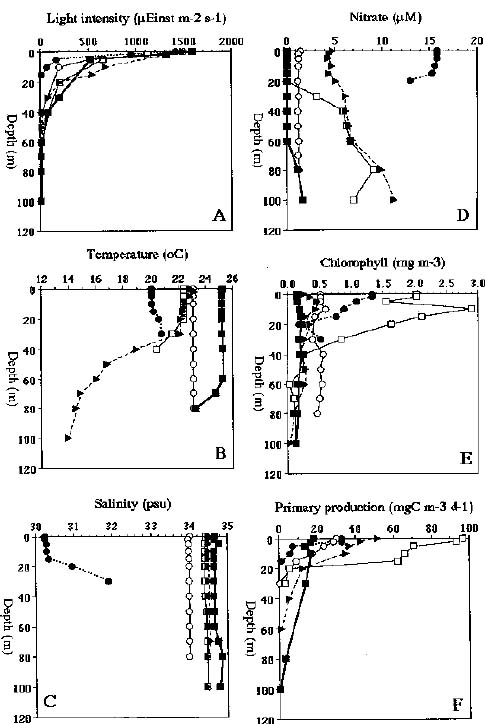
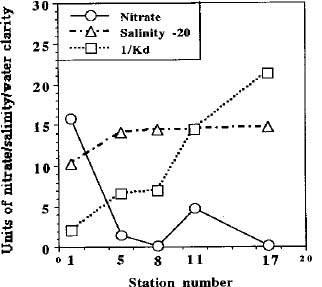
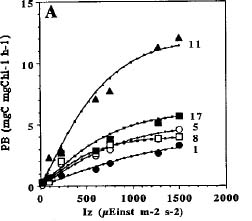
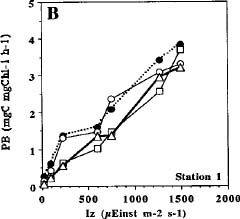
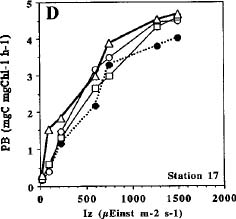
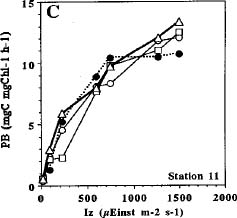
Abstract. Phytoplankton primary production was measured in different regions in the southern East China Sea north of Taiwan in the fall of 1994. Light-manipulation experiments were also performed on-board to study the photoacclimation of phytoplankton assemblages. The sampling stations encompassed the inner and outer shelf of the southern East China Sea. Field observation showed that the in situ maximal chlorophyll-normalized photosynthetic rates (PBmax) varied 3-fold among stations, ranging from 4.0 to 12.0 mgC mgChl-1 h-1. The low PBmax values recorded in the nutrient-laden coastal water and the oligotrophic Kuroshio water could be ascribed to high turbidity and low nutrient availability, respectively. The highest PBmax observed in the upwelling plume station probably could be ascribed to both copious nutrient supply by upwelling processes and higher light availability. Light-manipulation experiments showed that in the well-mixed coastal water, the light response curves for samples taken from different depths of the euphotic zone were very similar. In both the upwelling region and the Kuroshio water, phytoplankton assemblages living below the mixed-layer depth were shade-acclimated while those living above that depth were light-acclimated. Overall, our results indicate that photosynthetic available radiance, light history and nutrient availability are crucial in controlling the spatial variation of algal photosynthetic performance in the study area.
Keywords: Assimilation number; East China Sea; Kuroshio; Photoacclimation; Primary production; Upwelling.
Introduction
Studying the temporal and spatial variability of primary production and the factors that control them is essential in understanding the biogeochemical cycle of carbon in the ocean (Knauer, 1991; Longhurst and Harrison, 1989). The assimilation number (i.e. Pmax; mgC mgChl-1 h-1) at optimum (saturating) light intensity, has been a widely-accepted index for the study of phytoplankton physiology and ecology (for review, see Cullen et al., 1992). Light intensity has been recognized as the most basic factor in controlling Pmax.
Many models have been proposed to analyze the effect of light on photosynthesis (Frenette et al., 1993 and citations therein). One of the most extensively used models is that dealing with the chlorophyll-normalized photosynthesis rate vs. light intensity below the onset of photoinhibition (i.e. PB - I curve without photoinhibition term; Jassby and Platt, 1976). This model is expressed as: PB = (Pmax) × tanh (A × Iz/Pmax), where PB (mgC mgChl-1 h-1) is the chlorophyll normalized photosynthetic rate; Iz, the light intensity (mEinst m-2 s-1) at depths z and A (mgC mgChl-1 h-1 mEinst m-2 s-1), the initial slope of the PB - I curve. However, the PB - I curve and values of Pmax change with seasons and locations and even covary with other environmental factors such as nutrients and temperature. For
example, Pmax for marine phytoplankton varies more than 100-fold, ranging from 0.2 to 40.0 mgC mgChl-1 h-1 (Harrison and Platt, 1980). Therefore, study of the seasonal and temporal variations of the empirical PB - I curves is essential for choosing an appropriate model in a given area. Furthermore, these empirical PB - I curves are very important in developing bio-optical models which can then be combined with the ocean color remote sensing technique to estimate primary production from satellite images (Balch et al., 1989; Morel, 1991; Platt and Sathyendranath, 1993).
Unlike higher plants, marine phytoplankton can not remain fixed in space with respect to a light field. As a result of turbulence, phytoplankton cells may experience large variations in light intensity over the course of a day. Considering the light regimes experienced by phytoplankton in natural waters, first-order variations in the light regimes are influenced by the path of the sun (i.e. the diurnal photoperiod). Second-order variations are affected by the vertical displacement of cells in the water column. Many species of phytoplankton adjust their cellular chlorophyll content and modify their photosynthesis-irradiance curves in response to second-order variations in the light regimes. This phenomenon is known as photoacclimation or light-shade acclimation.
Primary production in the southern East China Sea and the Kuroshio areas has been studied by several workers (Shiah et al., 1995 and citations therein). However, avail
4Corresponding author. Fax: 886-2-363-2583.