Bot. Bull. Acad. Sin. (1996) 37: 165-171
Chang et al. RAPD for detection of variation in Phytophthora cinnamomi
Use of random amplified polymorphic DNA markers for the detection of genetic variation in Phytophthora cinnamomi in Taiwan
T. T. Chang1, W. W. Yang, and W. Y. Wang
Division of Forest Protection, Taiwan Forestry Research Institute, 53 Nan-Hai Road, Taipei, Taiwan, Republic of China
(Received November 10, 1995; Accepted May 1, 1996)
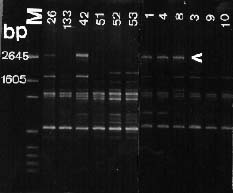
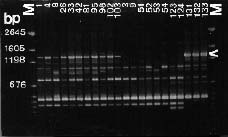
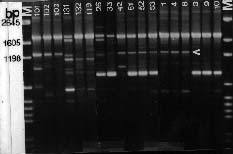
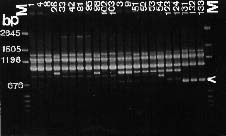
Abstract. Taiwanese isolates of Phytophthora cinnamomi were examined for genetic diversity by the random amplified polymorphic DNA assay. The amount of genetic variation was evaluated by polymerase chain reaction amplification with a set of nine random 10-mer primers. All amplifications revealed scorable polymorphisms among the isolates, and a total of 217 band positions was scored for the nine primers tested. Genetic distances between each of the isolates were calculated and cluster analysis was used to generate a dendrogram showing relationships between them. Isolates did not cluster into groups corresponding to their mating types. Primer OPS-13 did produce one 2.7 kb band for all isolates of A1 mating type, but not for A2 isolates. Moreover, primer OPE-06 amplified one 1.3 kb band for all A1 mating type isolates, while that was absent in A2 isolates except for three isolates obtained from avocado roots. Genetic differentiation of ten isolates obtained from the same location between two mating types was significantly high, indicating that no hybridization occurred between A1 and A2 mating types in that situation. Primer OPE-4 amplified a 680 bp fragment which was only observed in three isolates obtained from avocado roots. Also, genetic differentiation between isolates from avocado and those from other sources was significantly strong (distance between 0.262 and 0.412). These results suggest host specified races might occur in P. cinnamomi and the RAPD technique could be used to differentiate races within P. cinnamomi.
Keywords: Phytophthora cinnamomi; RAPD; Taiwan.
Introduction
Phytophthora cinnamomi Rands is an important soil-borne plant pathogen in agricultural, horticultural, and forest ecosystems with a world-wide distribution (Zentmyer, 1980). P. cinnamomi is widely distributed in natural forests on the island of Taiwan (Ann, 1984; Ann and Ko, 1985; Ko et al., 1978). Meanwhile, the fungus appears to coexist with many indigenous plant species and causes no disease to those plants (Ko et al., 1978). But, P. cinnamomi has brought disease to many cultivated fruit trees and plantations of two Cinnamomum species. (Ann, 1984; Chang, 1993)
In Taiwan, the native flora in the natural forest which was inhabitated by both A1 and A2 mating types of P. cinnamomi remained healthy. Moreover, considerable variation in biological characteristics was noted among isolates of both A1 and A2 mating types of P. cinnamomi isolated from Taiwan. These observations led to the proposition that Taiwan was an Asian origin of P. cinnamomi (Ko et al., 1978) in accord with Vavilov's concept of centre of origin (Vavilov, 1951). However, genetic variation has not been used to assess this proposition
or the differentiation between A1 and A2 mating types of P. cinnamomi isolated from Taiwan.
The random amplified polymorphic DNA (RAPD) markers generated with single primers of arbitrary nucleotide sequence have been shown useful in detecting intraspecific polymorphisms among fungi (Assigbetse et al., 1994; Crowhurst et al., 1991; Zimand et al., 1994). This technique can generate specific DNA fragments used for genome mapping, identification of isolates, and applications in molecular ecology (Hadrys et al., 1992). For plant pathogenic fungi, RAPD analysis can provide markers to differentiate races of F. solani f. sp. cucurbitae Snyder & Hansen (Crowhurst et al., 1991), F. oysporum f. sp. pisi (Schlecht.) Snyder & Hansen (Grajal-Martin et al., 1993), F. o. f. sp. vasinfectum (Assigbetse et al., 1994), aggressive and nonaggressive isolates of Phoma lingam (Tode ex Fr.) Desm. (Goodwin and Annis, 1991; Schäfer and Wöstemeyer, 1992), isolates with different geographic origins of Collectotrichum graminicola (Ces.) Wils. (Guthrie et al., 1992) and isolates of Discula umbrinella Sacc. from different hosts (Haemmerli et al., 1992). This RAPD techique has led us to investigate the genetic diversity within P. cinnamomi. The objective of this study was to examine the relationships among 26 isolates of both mating types from Taiwan.
1Corresponding author. Fax: (02) 389-1419.