Bot. Bull. Acad. Sin. (1996) 37: 181-190
Yeh and Chu PWV coat protein transgenic tobacco
Production and evaluation of transgenic tobacco plants expressing the coat protein gene of passionfruit woodiness virus
Shyi-Dong Yeh1 and Fang-Hua Chu
Department of Plant Pathology, National Chung Hsing University, Taichung, Taiwan, Republic of China
(Received January 17, 1996; Accepted May 1, 1996)
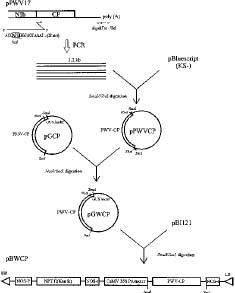
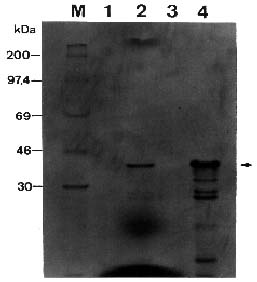
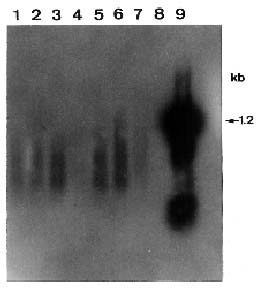
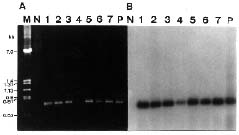
Abstract. Passionfruit woodiness virus (PWV), a member of the plant potyvirus group, is the major limiting factor for growing passionfruit in Taiwan. The coding sequence of the coat protein (CP) gene of PWV was modified to have an initiation codon and a leader sequence for translation. Analyses by in vitro transcription, in vitro translation, and immunoprecipitation verified that the modified PWV CP gene correctly encoded and expressed a protein of 36 kDa reacting to PWV CP antiserum. The CP gene was further inserted in a Ti binary vector and transferred to tobacco Nicotiana benthamiana by Agrobacterium-mediated transformation. Analyses of the putative transgenic plants by PCR amplification coupled with Southern blotting revealed an expected DNA fragment of 0.7 kb reacting with the probe specific to the PWV CP gene. Western blotting and Northern blotting were also used to further detect the expression of the PWV CP gene. The transgene-derived PWV CP was not detected but the degraded forms of the CP transcript were present. The putative transgenic tobacco lines of w-6, 9, 10, 12, 14, 15, and 16 were subjected to inoculation with different concentrations of PWV prepared from PWV-infected tobacco leaves. All plants of the four lines w-9, 10, 14, and 15 remained symptomless up to six weeks after inoculation. Some plants of two lines (w-6 and 9) showed a delay of 3 to 10 days in symptom development. All plants of line w-12 showed symptoms at the same time as the controls. Inoculated plants of lines w-9, 10, 14, and 15 were further tested by ELISA and bioassays on the local lesion host Chenopodium quinoa and the systemic host N. benthamiana. The negative results indicated that the complete resistance was provided by the apparent inhibition of PWV replication.
Keywords: Passionfruit woodiness virus; Transgenic tobacco plants; Coat protein gene; Potyvirus.
Introduction
Passionfruit woodiness virus (PWV), first described by McKnight (1953), is a potyvirus species, with flexuous particles of 750 × 12 nm and a genome consisting of a single stranded RNA of positive polarity (Taylor and Greber, 1973; Hollings and Brunt, 1981). PWV has been reported to infect plants of 44 species in 21 genera of 6 families (Edwardson and Christie, 1991). The virus is transmitted by aphids in a nonpersistent manner (Siaw, 1971) and mechanically by grafting or by contaminated cutting tools (Nattrass, 1944; Wang, 1983). The passionfruit woodiness disease caused by PWV is characterized by foliar symptoms of mosaic, distortion and rugosity, and the production of woody and severely malformed fruits (Taylor and Greber, 1973; Wang, 1983). Another feature of PWV infection is that the virus induces characteristic pinwheel and tubular inclusion bodies in the cytoplasm of infected cells (Moghal and Francki, 1981; Pares and McGechan, 1975; Lin et al., 1987; Jan and Yeh, 1995).
An isolate of PWV was first recorded in Taiwan in 1981 (Chang et al., 1981) and since then it has widely spread in the passionfruit-growing areas across the island as a major limiting factor on the production of the passionfruit
cultivar Tainung No. 1 (TN-1; a hybrid of Passiflora edulis Sims × P. edulis f. flavicarpa Degener). The virus has a single type coat protein (CP) of 36 kDa and produces a cylindrical inclusion protein (CIP) of 66 kDa and an amorphous inclusion protein (AIP) of 51 kDa in the cytoplasm of infected cells (Jan and Yeh, 1995). Antisera produced against the CP or virions of PWV have been widely used for detection of the virus by serological means (Lin et al., 1988; Chang, 1992). Since passionfruit in Taiwan is commercially propagated by grafting, indexing the presence of PWV in scions is important for the production of virus-free seedlings.
Plant viruses cause significant losses to crops worldwide and the basic approaches for controlling them have not been overly successful. The new concept of nonconventional resistance, which involves transforming plants with nucleic acid sequences that interfere with the viral infection cycle, is a promising new approach. Since the initial demonstration that expression of the CP gene of tobacco mosaic virus (TMV) in transgenic tobacco could provide effective resistance against TMV infection (Powell-Abel et al., 1986), genetically engineered CP-mediated protection has been used with success against several virus groups (for reviews, see Beachy et al., 1990; Hull and Davies, 1992; Fitchen and Beachy, 1993). In each case, expression of a viral CP gene conferred protection against different strains of the same virus and the resistance was nuclearly inherited.
1Corresponding author. Fax: 04-2877585; E-mail: sdyeh@ nchu.edu.tw.