Bot. Bull. Acad. Sin. (1996) 37: 213-218
Hannweg et al. Micropropagation of Bowiea volubilis
A simple method for the micropropagation of Bowiea volubilis from inflorescence explants
K. Hannweg, M.P. Watt1, and P. Berjak
Centre for Indigenous Plant Use Research, Biology Department, University of Natal, Private Bag X10, Dalbridge 4014, Republic of South Africa
(Received February 26, 1996; Accepted June 17, 1996)
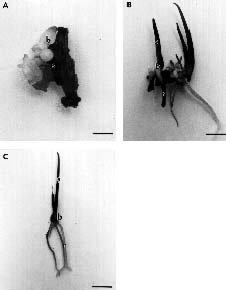
Abstract. The bulb of Bowiea volubilis (Liliaceae) is important in traditional medicine in South Africa. Although this species can be macro- and micropropagated, present methods relying on the use of bulb-scale pieces can result in the destruction of the parent plant. In this study, a protocol for the organogenic plant regeneration from inflorescence pieces was established. It involves the culture of 10 mm long pieces of inflorescence stem on MS basal nutrient medium, 30 g l-1 sucrose, 10 g l-1 agar, 1 mg l-1 2,4-D and 1 mg l-1 BAP, in the dark for 6_8 weeks. The explants are then transferred to fresh medium without growth regulators, where bulblet development, shoot elongation and rooting occur within 4_5 weeks. Successful hardening-off is achieved using conditions of relatively low humidty. As the inflorescence is very long (up to 20_30 m in total) and the protocol results in approximately 4.6 plantlets per explant, thousands of plantlets can potentially be produced in this manner from a single inflorescence.
Keywords: Conservation; Ethnobotany; Medicinal plant; Propagation.
Introduction
Bowiea volubilis (Zulu name = igibisila), a member of the Liliaceae, is a geophyte endemic to the central and eastern regions of South Africa (Dyer, 1964). The bulb is globose, up to 150 mm in diameter and may be subterranean or exposed. The leaves are lanceolate, canaliculate, up to 350 mm long and 5 mm in diameter. The stem producing the annual inflorescence can be up to 3 m long and appears during the summer months.
Although the bulb of B. volubilis has long been known to be poisonous (Watt and Breyer-Brandwijk, 1962), it forms an important and very popular component of traditional medicine in South Africa. It has been reported to contain three active cardiac glucosides, one of which appears to act as a heart stimulant (Watt and Breyer-Brandwijk, 1962). It is alleged to be of medicinal value and preparations of the bulb alone or mixed with parts of other species are used widely as a purgative, a remedy for dropsy and female infertility, and as components of skin and eye lotions. The bulb is also purported to have magical properties. It is sprinkled on impis' to ward off enemies and is used as an aphrodisiac (Watt and Breyer-Brandwijk, 1962; Mkhize, pers. comm.).
At present, the demand for Boweia volubilis is satisfied primarily from remaining wild populations. However, its popularity as an herbal medicine coupled with the rapid human population expansion and dramatic decrease in the areas of indigenous vegetation have led to the
overexploitation and depletion of wild stocks and consequent rapid increases in prices. In 1990, the sale value of Bowiea volubilis was $5.10 per dozen plants, and over two tons of bulb material were sold each year (Buckas, pers. comm.). It is not suprising, therefore, that even eight years ago B. volubilis was one of the top five medicinal species nominated by herb traders as having become scarce (Cunningham, 1988).
Very little is known about the natural propagation of B. volubilis. However, it has been reported that few seeds are set (Dyer, 1964) and hence progeny numbers are low. Traditional methods of vegetative propagation of bulbs such as daughter-bulb formation have been attempted in nurseries, but these techniques provide an extremely slow rate of multiplication. Thus, there has been a distinct need to investigate the potential of in vitro propagation systems for this plant, as has been successfully achieved with other bulbous species of commercial value (e.g. Allium spp.: Havraneck and Novak, 1976; Amaranthus spp.: Flores et al., 1981).
Bowiea volubilis has been previously propagated using tissue culture techniques. Jha and Sen (1986) used a shake culture system and Cook et al. (1988) successfully produced B. volubilis in solid culture using bulb scale material as explants. However, the removal of the inner bulb scales (which are the youngest, least contaminated, and most responsive to culture) results in the destruction of the parent plant, a great disadvantage in terms of conservation of an endangered species. In this study, we investigated the feasibility of establishing a protocol for the micropropagation of B. volubilis from inflorescence explants.
1 Corresponding author. Fax: 27-31-2602029/1195; Email: watt@biology.und.ac.za.