Bot. Bull. Acad. Sin. (1996) 37(4): 239-245
Lee and Lin Rice coleoptile peroxidase
Peroxidase activity in relation to ethylene-induced rice (Oryza sativa L.) coleoptile elongation
Tse-Min Lee1 and Yaw-Huei Lin2,3
1Institute of Marine Biology, National Sun Yat-Sen University, Kaohsiung, Taiwan, Republic of China
2Institute of Botany, Academia Sinica, Taipei, Taiwan, Republic of China
(Received April 1, 1996; Accepted June 19, 1996)
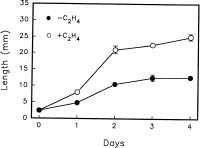
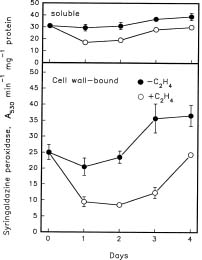
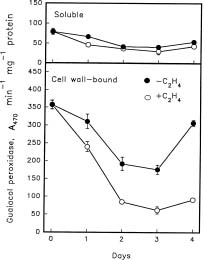
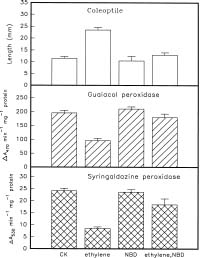
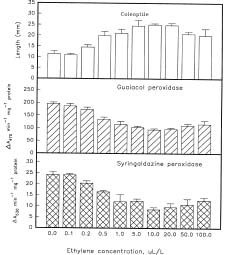
Abstract. Changes in the activities of soluble and cell wall-bound peroxidase (EC 1.11.1.7, donor: hydrogen-peroxide oxidoreductase) in relation to the regulation of coleoptile elongation were determined using rice (Oryza sativa L. cv. Taichung Native 1) seedlings grown in air or in air containing ethylene. Guaiacol and syringaldazine were used separately as the H donors for peroxidase assays. In air, both guaiacol and syringaldazine cell wall-bound peroxidase activity declined during coleoptile elongation but increased again to the initial level when elongation ceased. Soluble syringaldazine peroxidase activity in air-grown coleoptiles also showed an initial decline followed by a subsequent rise, but soluble guaiacol peroxidase activity remained essentially constant. Ethylene (10 µL/L) enhanced coleoptile elongation but decreased both guaiacol and syringaldazine peroxidase activities, especially the wall-bound forms. 2,5-Norbornadiene (NBD) at 3000 µL/L, an inhibitor of ethylene action, blocked both the ethylene-mediated coleoptile elongation and the decline of peroxidase activity. Ethylene at 50 µL/L reversed the effect of NBD. These results suggest that the activity of peroxidase, especially the cell wall-bound form, is inversely related to the elongation of rice coleoptiles.
Keywords: Oryza sativa L.; Coleoptile elongation; Ethylene; Peroxidase; 2,5-Norbornadiene.
Abbreviation: NBD, 2,5-Norbornadiene.
Introduction
Rice is one of the plants that can germinate and grow in anoxia or submergence conditions (Alpi and Beevers, 1983; Ishizawa and Esashi, 1984; Raskin and Kende, 1983; Taylor, 1942; Turner et al., 1981). However, the growth of rice seedlings under oxygen-restricted environments is limited to coleoptile elongation (Atwell et al., 1982; Turner et al., 1981). This rapid coleoptile elongation is considered to be a survival mechanism for rice seedlings to allow them to grow out of flood water and obtain oxygen.
Ethylene enhances rice coleoptile elongation (Ishizawa and Esashi, 1984; Ku et al., 1970; Lee and Chu, 1992; Raskin and Kende, 1983; Salter and Kende, 1985) with several physiological factors involved. One is osmoregulation by a stimulated sucrose transport from endosperm to coleoptiles (Ishizawa and Esashi, 1988). Another factor is enhanced cell wall loosening by alteration of wall components. For example, the decline of the level of (1,3), (1,4)-b-D-glucans has been shown to be closely associated with the ethylene-induced cell wall loosening in rice coleoptiles (Hoson et al., 1990).
Peroxidase (EC 1.11.1.7, donor: hydrogen-peroxide oxidoreductase), which is a glycoprotein, has been recognized as mediating the binding of ferulic acid to cell walls by the formation of diferuloyl cross-links to matrix po
lysaccharides, and its activity subsequently reduces wall extensibility (Fry, 1979). Peroxidases are also responsible for the assembly of lignins and proteins in cell wall (Fry, 1986; Gaspar et al., 1985; van Huystee, 1987; Iiyama et al., 1994; Taiz, 1984; Whitmore, 1978). Higher peroxidase activities are closely associated with reduced growth of plants including peanut hypocotyls (Zheng and van Huystee, 1992), mung bean hypocotyls (Goldberg et al., 1987) and dwarf sorghum (Schertz et al., 1971). Results from two genotypes of tall fescue (Festuca arundinacea Schreb.) with different lengths of leaf elongation zones show that the cessation of elongation is accompanied by an increase of cell wall-bound peroxidase activity (MacAdam et al., 1992).
The activity of peroxidase, especially the cell wall-bound form, is related to ethylene-regulated plant growth. Petioles of Ranunculus sceleratus L. elongate faster and have lower peroxidase activities in response to ethylene (Horton, 1993). In contrast, the ethylene-induced inhibition of pea epicotyl elongation is accompanied by a rise of peroxidase activity (Ridge and Osborne, 1970). In the case of the inversion-induced growth inhibition of Pharbitis shoots, ethylene levels increase during inversion, and the subsequent increase of wall-bound peroxidase activity is correlated with wall stiffening (Prasad and Cline, 1987). There is also a negative correlation between coleoptile elongation and cell wall-bound peroxidase levels in anoxia-treated rice seedlings (Lee and Lin, 1995).
3Corresponding author. Fax: 886-2-7827954.