Bot. Bull. Acad. Sin. (1996) 37(4): 247-253
Liu et al. Effect of NAA on indoleacetic acid in hypocotyl cuttings of soybean during root formation
Effect of naphthaleneacetic acid on endogenous indole-3-acetic acid, peroxidase and auxin oxidase in hypocotyl cuttings of soybean during root formation
Zin-Huang Liu1,3, I-Chao Hsiao2, and Ya-Wen Pan2
1Department of Biology, Institute of Life Sciences, National Sun Yat-Sen University, Kaohsiung, Taiwan 804, Republic of China
2Department of Biology, Tunghai University, Taichung, Taiwan, Republic of China
(Received April 8, 1996; Accepted August 16, 1996)
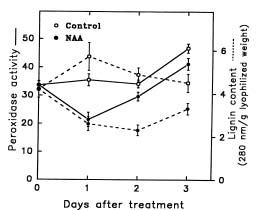
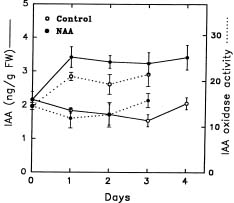
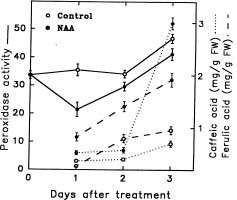
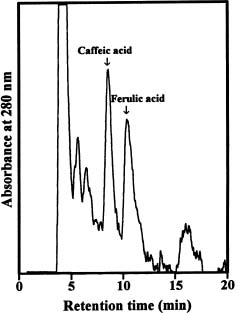
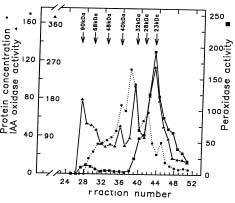
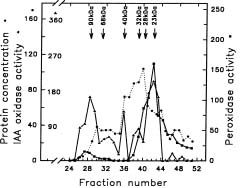
Abstract. The effect of exogenous naphthaleneacetic acid (NAA) on the internal levels of indole-3-acetic acid (IAA) in rooting hypocotyls of Glycine max was studied. The hypocotyls of NAA-treated cuttings grew significantly higher numbers of adventitious roots with an increase in endogenous IAA levels that corresponded with a decrease in IAA oxidase activity (32 kDa) examined. Moreover, a decline of peroxidase activity was accompanied by a decrease of lignin content during root formation. Caffeic acid and ferulic acid, two critical phenolic compounds for lignin synthesis, accumulated in NAA-treated tissues. Consequently, the increased IAA levels with a decrease of IAA oxidase activity accompanied a lower lignin content and a reduced peroxidase activity in NAA-treated tissues suggests that the induction of adventitious roots by NAA in soybean cuttings may be due to to the higher IAA levels accumulated in tissues.
Keywords: Adventitious rooting; Indole-3-acetic acid; IAA oxidase; Naphthaleneacetic acid; Peroxidase; Glycine max.
Abbreviations: HPLC, high performance liquid chromatography; IAA, indole-3-acetic acid; NAA, naphthaleneacetic acid; TLC, thin layer chromatography.
Introduction
It is known that adventitious root formation can be stimulated by exogenously applied auxins, yet the mechanism of this physiological response is still disputed. Conflicting results have been reported about the endogenous indole-3-acetic acid (IAA) levels in rooting tissues. In most cases, high levels of IAA are associated with the promotion of adventitious rooting (Weigel et al., 1984; Moncousin et al., 1989; Liu and Reid, 1992; Sagee et al., 1992). In some cases, the levels of endogenous IAA declined during root formation (Hausman, 1993). Furthermore, some reports have indicated that root initiation occurred without any changes of IAA levels in the root regenerating zone (Nordstrom and Eliasson, 1991). The regulation of plant development by IAA may depend on the amount of free IAA present in plant tissues. Free IAA can be modulated via several pathways, including IAA metabolism, synthesis and transport, and the breakdown of conjugated IAA. One well known IAA metabolic pathway consists of the oxidative decarboxylation of the side chain of IAA by IAA oxidase, leading to the formation of either indole-3-methanol or 3-methyleneoxindole (Grambow and Langenbeck-Schwich, 1983; Beffa et al., 1990). Moreover, numerous papers have reported that
auxin-induced changes in peroxidase and IAA oxidase occur during the rooting processes (Gaspar et al., 1985; Mato et al., 1988; Fett-Neto et al., 1992). Yet, little is known about the correlation between IAA levels and IAA oxidase activity during the processes of rooting. One aim of this investigation is to examine the relationship between the change of IAA content and IAA oxidase activity during root formation.
The formation of adventitious roots involves the process of redifferentiation, in which predetermined cells switch from their morphogenetic path to act as mother cells for the root primordia (Friedman et al., 1979; Aeschbacher et al., 1994). Among these changes, the process of lignification in the cell wall, catalyzed by a particular peroxidase, may occur during the rooting (Fukuda and Komamine, 1982; Church and Galston, 1988; Bruce and West, 1989; McDougall, 1992; Sato et al., 1993). In addition to examining the changes in IAA content and IAA oxidase activity, the effect of NAA on lignification during the rooting processes is also investigated.
Materials and Methods
Plant Materials and Rooting Experiments
Soybean (Glycine max) seedlings were germinated and grown in vermiculite at 27 ± 0.5°C under 16 h/8 h light/
3Corresponding author. Fax: 886-07-5319333.