Bot. Bull. Acad. Sin. (1997) 38: 21_28
Hwang and Chen Ultrastructure of Kandelia candel
Effect of tonicity and additives to the fixative on ultrastructure of mesophyllous cells in Kandelia candel (L.) Druce (Rhizophoraceae)
Yuan-Hsun Hwang1 and Shuh-Chun Chen
Institute of Botany, Academia Sinica, Taipei, Taiwan 115, Republic of China
(Received May 24, 1996; Accepted August 8, 1996)
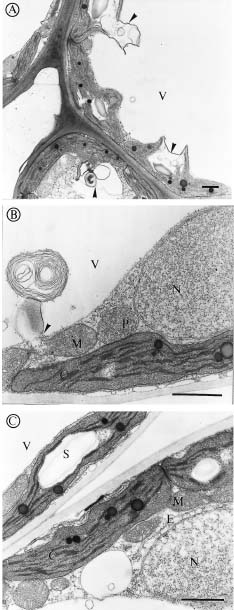
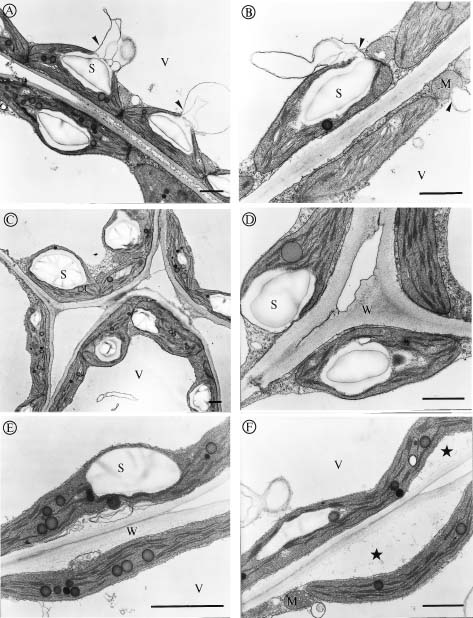
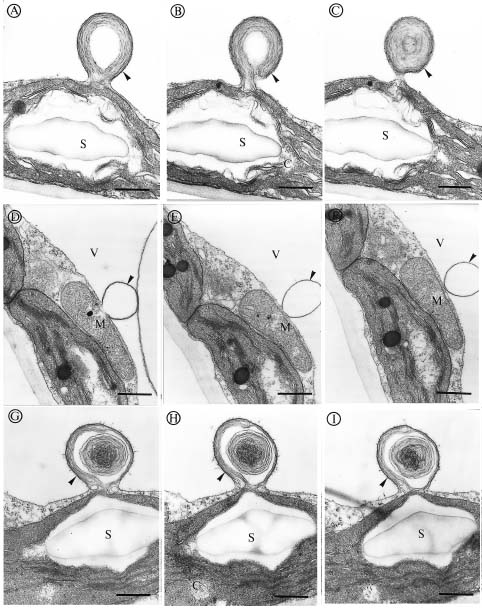
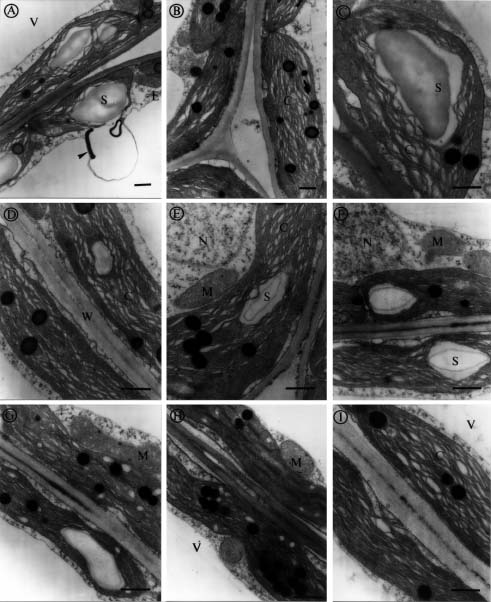
Abstract. We investigated the effect of tonicity and additives to the fixative on the ultrastructure of Kandelia candel. Nonionic additives such as sucrose were better than ionic additives such as NaCl, KCl, Na2SO4 or K2SO4. Measurement of osmolality in the K. candel cells was carried out for adjusting the osmolality of the fixatives. We found that the hypertonic fixative and the washing buffer obtained by adding 0.15 M sucrose to the isotonic fixative and 0.1 M phosphate buffer yielded the best results; outer membranes of chloroplasts and mitochondria did not dilate or undergo plasmolysis. Varying the tonicity of the fixative with sucrose to a reasonable degree did not change the ultrastructural details within the chloroplasts and mitochondria. In contrast, hypertonic fixatives obtained by adding ionic additives caused significant swelling of thylakoid membranes in chloroplasts. This artifact was most pronounced when hypertonic fixatives containing NaCl or KCl were used. These observations suggest that high ionic fixatives disturb the ionic balance in thylakoid membranes.
Keywords: Kandelia candel; Mangrove; Osmotic potential; Salt; Sucrose; Tonicity.
Abbreviations: Ys, Osmotic potential; FB, Fixative and washing buffer; LSP, 100 mM NaCl treated plant; HSP, 400 mM NaCl treated plant.
Introduction
Salinity has been known to affect the ultrastructure of plants, both glycophytes and halophytes. The swelling of organelles such as, chloroplasts, golgi bodies, mitochondria, and nuclei is probably the most obvious phenomenon found in plants growing in high salinities, which was suggested as a response to the changing internal environment of plant tissues (Blumenthal-Goldschmidt and Poljakoff-Mayber, 1968; Poljakoff-Mayber, 1975; Werker et al., 1983). Other studies found increasing formation of vesicles and myelin-like structures in the vacuoles of plants growing in high salinities, and these structures were thought to be the pinocytotic invagination of tonoplast, which allows plants to translocate salt ions from the cytoplasm into the vacuoles (Willert and Kramer, 1972; Kurkova and Balnokin, 1994).
However, most of the above studies using fixatives for electron microscopy did not mention the relationship between the tonicity of the fixative and the osmolality of the specimen. The effect of tonicity of fixatives on the ultrastructure has been demonstrated mostly in animal cells (Schultz and Karlsson, 1965; Maunsbach, 1966; Bone and Denton, 1971; Rasmussen, 1974); however, a few studies were carried out in plant systems (Fineran, 1971; Soikkeli,
1980). Although some of the studies mentioned above used either sucrose (Werker et al., 1983) or NaCl (Kurkova and Balnokin, 1994) in the fixative and buffer to maintain an osmolality equivalent to that of the culture medium of the plant cells, the effect of the tonicity of the fixative on the ultrastructure of the plant samples was not described.
Kandelia candel, a mangrove, like other halophytes (Flowers et al., 1977), were used to accumulate high salt ions to reduce the osmotic potential in tissues even growing in normal Hoagland's solution (Hwang and Chen, 1995). Moreover, the osmotic potential in root and leaves of K. candel was correlated with, and lower than, the osmotic potential of the saline culture solutions (Hwang and Chen, 1995). We noticed that the outer shape of organelles was affected tremendously by the tonicity of fixatives while we were studying the ultrastructure of K. candel. Therefore, the present investigation was carried out to determine how the ultrastructure of leaf cells in K. candel grown in varying NaCl salinities were influenced by the tonicity and different additives in the fixatives.
Materials and Methods
Plant
Mature propagules of Kandelia candel (L.) Druce, collected from mature trees growing along the Tamshui River, Taipei, Taiwan, (121°26'E, 25°9'N), were cultivated on sand in pots, partially submerged in nutrient solutions. The
1Corresponding authors. Fax: 886-2-7827954; E-mail: bohwang @ccvax.sinica.edu.tw