Bot. Bull. Acad. Sin. (1997) 38: 63_72
Wu New Hyphoderma from Taiwan
New species and new records of Hyphoderma (Basidiomycotina) from Taiwan
Sheng-Hua Wu1
Division of Collection and Research, National Museum of Natural Science, Taichung, Taiwan, Republic of China
(Received September 9, 1996; Accepted November 27, 1996)
Abstract. Four new species of Hyphoderma collected from Taiwan are presented, viz. H. clavatum, H. rimulosum, H. subclavatum, and H. subpraetermissum. Two species are additionally reported from Taiwan for the first time, viz. H. definitum and H. variolosum. A key to known species of Taiwanese Hyphoderma is given. Description, microscopic line drawings, sexuality, cultural characters, and nuclear behavior are presented for the four new species.
Keywords: Basidiomycotina; Cultural studies; Hyphoderma; Taiwan; Taxonomy.
Introduction
Hyphoderma Wallr. is usually treated by mycologists under the heterogeneous Corticiaceae Herter (Donk, 1964; Eriksson and Ryvarden, 1975; Tellería, 1990; Wu, 1990). More than ninety species of Hyphoderma are known (e.g., Hjortstam, 1987; Hjortstam and Larsson, 1994; Wu, 1997), representing the largest genus among the Corticiaceae s.l.
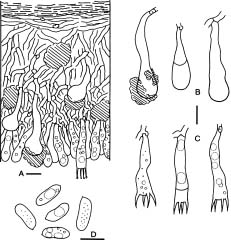
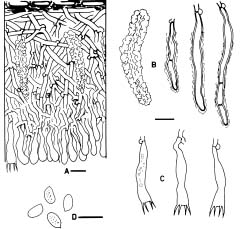
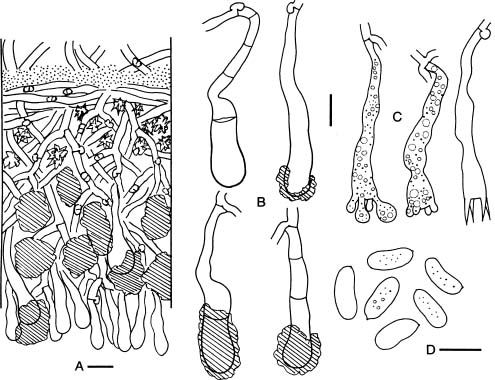
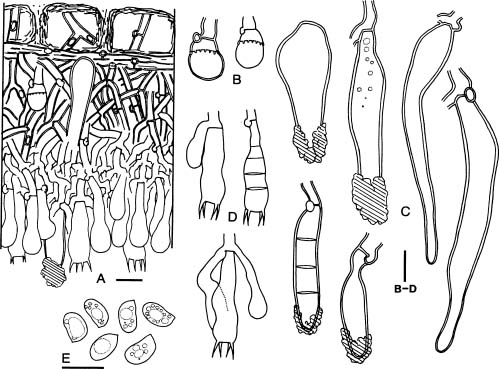
Basidiomata of Hyphoderma are resupinate and effuse. Most species have smooth hymenial surfaces, but tuberculate, grandinioid, or odontioid cases are present in some species. Hyphoderma is microscopically characterized by the subclavate basidia with a median constriction, and typically monomitic hyphal system and nodose-septate generative hyphae, as well as nonamyloid and acyanophilic basidiospores, which are basically thin-walled. Leptocystidia, encrusted cystidia, or both are present in most species. Sexually, most species of Hyphoderma have been reported as heterothallic bipolar. Hyphoderma is lignicolous, saprobic, and causes a uniform white rot in wood. Tzean and Liou (1993) have shown that Hyphoderma is the only one among several tested genera of resupinate basidiomycetes that can capture or poison and consume nematodes.
Lin and Chen (1990) first reported Hyphoderma species from Taiwan, viz. H. praetermissum (P. Karst.) J. Erikss. & Strid, H. rude (Bres.) Hjortstam & Ryvarden (as Hyphodontia mucronata (Furukawa) S.H. Lin & Z.C. Chen), and H. setigerum (Fr.) Donk. Nine Hyphoderma species were further presented by Wu (1990) as new to Taiwan: H. allantosporum Sheng H. Wu, H. argillaceum (Bres.) Donk, H. ayresii (Berk. ex Cooke) Hjortstam (as H. macrosporum Sheng H. Wu), H. hjortstamii Sheng H. Wu, H. litschaueri (Burt) J. Erikss. & Strid, H. malenconii (Manjón & Moreno) Manjón et al., H. microcystidium
Sheng H. Wu, H. neopuberum Sheng H. Wu and, H. puberum (Fr.:Fr.) Wallr. Recently, Wu (1997) reported four additional new species of Hyphoderma from Taiwan: H. acystidiatum Sheng H. Wu, H. cremeum Sheng H. Wu, H. densum Sheng H. Wu, and H. subsetigerum Sheng H. Wu. However, the survey of Hyphoderma in Taiwan is still incomplete. In this study, four new species and two new records of Hyphoderma are proposed from Taiwan. Cultural studies including sexuality and nuclear behavior are also provided for the four new species.
Materials and Methods
Materials for this study were collected from Taiwan by the author in recent years. All studied specimens and cultures are deposited at the herbarium of National Museum of Natural Science of ROC (TNM).
Free-hand thin sections of basidiomata were prepared for microscopic studies. For observations and measurements of microscopic characters, 5% KOH was used as mounting medium to ensure rehydration. Melzer's reagent (IKI) was employed to detect amyloidity and dextrinoidity. Cotton blue (CB) was used as a mounting medium to determine cyanophily. Cultural description and species code system are basically from Nobles (1965) with amendments by Boidin and Lanquetin (1983). Minor modifications to Nobles' code system have been presented by many mycologists. The Nobles' code detailed by Nakasone (1990) is comprehensive and adopted in this study. Following Nakasone (1990) the mycelia were grown on 1.5% MEA instead of 1.25% MEA. In this study, plates were inverted to avoid accumulation of water produced by the mycelia. Inverted plates permit formation of a hymenium oriented as in nature. Nuclear staining of mycelia were made with Giemsa according to Boidin (1958). DAPI (4'-6'-diamidino-2-phenylindole) was used at a concentration of 0.25 µm/ml as fluorescent stain for nuclei of basidiospores.
1Fax: +886-4-323-5367; Email: shwu@nmns1.nmns.edu.tw