Bot. Bull. Acad. Sin. (1997) 38: 177-182
Zheng and Ko Variation of Phytophthora cinnamomi
Continuing variation in colony morphology and fungicide
sensitivity in Phytophthora cinnamomi following exposure
to chloroneb
Xia Bo Zheng1,2 and Wen-Hsiung Ko1,3
1Department of Plant Pathology, Beaumont Agricultural Research Center, University of Hawaii at Manoa, Hilo, Hawaii 96720, U. S. A.
(Received December 9, 1996; Accepted March 8, 1997)
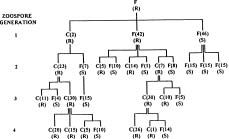
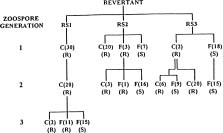
Abstract. A homothallic isolate of Phytophthora cinnamomi grown on agar medium containing fungicide chloroneb, produced fast growing resistant sectors within 3 weeks. The colony of a fungicide-resistant mutant, which was similar to the original flat (F) type, gave rise to single zoospore cultures consisting of a new cottony (C) type resistant to chloroneb and the F type either resistant or sensitive to chloroneb. The F type was continuously segregated from the C type in three successive zoospore generations. Although the chloroneb-sensitive F type revertants were stable, they produced more resistant sectors in a shorter time than did wild type when exposed again to chloroneb. Single zoospore cultures obtained from these resistant sectors were C type resistant to chloroneb and F type either resistant or sensitive to chloroneb. Both C and F type cultures resistant to chloroneb were unstable and segregated in successive zoospore generations in a fashion similar to that described above. The majority of zoospores (89 to 99%) produced by both C and F types were uninucleate.
Keywords: Chloroneb; Colony morphology; Fungicide sensitivity; Phytophthora cinnamomi; Variation.
Introduction
Phytophthora cinnamomi Rands usually produces clusters of chlamydospores, coralloid hyphae, and both spherical and irregular hyphal swellings on agar media, and forms uniform nonpapillate sporangia only under special conditions such as in nonsterile soil extract (Zentmyer, 1980). The fungus is also considered heterothallic because of the requirement for the presence of opposite mating types for production of sexual structures (Ho, 1981; Newhook et al., 1978). However some isolates of P. cinnamomi vary from the characteristic features mentioned above. Ann and Ko (1985) reported the isolation of an A2 isolate of P. cinnamomi from soil capable of producing sporangia on agar medium and A1 isolates from citrus incapable of producing chlamydospores. From black locust (Robinia pseudoacacia L.) Ho et al. (1983) obtained an A1 isolate of P. cinnamomi incapable of producing chlamydospores, but capable of producing sporangia on agar medium, and a homothallic isolate incapable of producing chlamydospores.
During the selection of fungicide resistance as a marker for studying the genetic relationship between the homothallic isolates and the heterothallic isolates of P.
cinnamomi, it was found that exposure of the fungus to chloroneb not only induced resistance to this fungicide but also altered the colony morphology. Fungi usually are inhibited by fungicides only in the presence of those chemicals, and revert to normal growth and appearance after being transferred to a medium free from these chemicals. Although development of resistance to fungicides by fungi after their exposure to these chemicals is relatively common (De Waard et al., 1993), reports on changes in other characteristics induced by fungicides are rare. We, therefore, studied the stability of the new colony type and the relationship between the new colony type and the fungicide resistance induced by chloroneb. Details of the study are reported here.
Materials and Methods
Microorganism
Isolate P630 of P. cinnamomi was obtained from a diseased tree of black locust in Jiangsu, China and was derived from a single zoospore. The fungus was homothallic, capable of producing oospores in single culture, but incapable of forming chlamydospores (Ho et al., 1983). It was also distinctive in having the ability to produce sporangia in diluted sterile soil extract.
Induction of Sector Formation
The method described by Ann and Ko (1985) was used to induce sector formation. Four culture blocks (ca.
2Present address: Department of Plant Protection, Nanjing Agricultural University, Nanjing, China.
3Corresponding author. Fax: (808) 969-7923.