Bot. Bull. Acad. Sin. (1998) 39: 29_32
Shih and Kao Acid phosphatase under stress condition
Induction of acid phosphatase in detached rice leaves under stress conditions
Chiung-Yueh Shih and Ching Huei Kao1
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
(Received March 13, 1997; Accepted September 9, 1997)
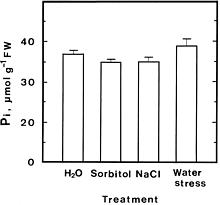
Abstract. The effect of water stress, sorbitol and NaCl on acid phosphatase (APase) activity in detached rice leaves was investigated. APase activity was increased in detached rice leaves treated with methyl jasmonate (MJ), water stress, sorbitol, and NaCl. The induction of APase activity under water, osmotic and salt stresses, and MJ was not accompanied by a decrease in phosphorus level. These results suggest that the induction of APase under stress conditions and MJ may have a cause other than phosphorus deficiency. Evidence is presented to show that stress-induced APase activity is due to de novo APase synthesis and is unlikely to be mediated through MJ.
Keywords: Acid phosphatase; Methyl jasmonate; Oryza sativa; Osmotic stress; Salt stress; Water stress.
Introduction
Acid phosphatase (APase, EC 3.1.3.2) is widely distributed in plants. It has long been recognized that APase activity in plants typically increases when plants become phosphorus (Pi) deficient. The increase in APase activity correlates with a low level of Pi in numerous species and plant parts (Barrett-Lennard et al., 1982; Duff et al., 1989; Goldstein et al., 1989; Lefebvre et al., 1990; Ueki and Sato, 1977). Salt, water, and osmotic stresses have also been reported to increase APase activity (Barrett-Lennard et al., 1982; Pan, 1987; Szabo-Nagy et al., 1992). It has been demonstrated that drought induced an increase in APase activity without exerting significant changes in the Pi level (Barrett-Lennard et al., 1982). Szabo-Nagy et al. (1992) also demonstrated that the induction of APase under osmotic and salt stresses was not accompanied by a decrease in Pi level. Recently, we reported that methyl jasmonate (MJ) markedly induced APase activity in detached rice leaves under light conditions and this induction is not caused by a decrease in Pi level (Yeh et al., 1995). It is not known whether the mechanism of APase activity induced by water, salt, and osmotic stresses is similar to that of MJ.
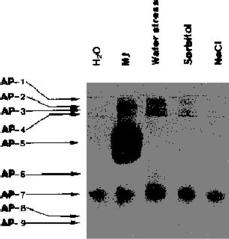
This study was designed to examine whether a low level of Pi can serve as a signal for the induction of APase under water, salt, and osmotic stresses in detached rice leaves. We also compared the effect of MJ and stresses on the isoenzymes of APase in detached rice leaves.
Materials and Methods
Rice (Oryza sativa, cv. Taichung Native 1) seedlings were grown in hydroponic culture as described previously
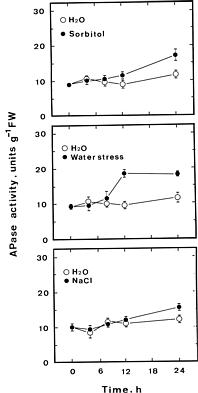
(Chen et al., 1990). The apical 3 cm of the third leaves of 12-day-old seedlings were used for the experiments. Ten segments of rice leaves were floated on 10 ml of test solution in a Petri dish. Incubation was conducted under light conditions (40 µmol m-2 s-1). Water stress was applied by a previously described method (Kao, 1981). Groups of ten leaf segments were weighed and exposed to the vapor above a solution of 0.5 M NaCl.
For enzyme extraction, leaf segments were homogenized in a prechilled mortar and pestle with cold 50 mM Tris-Mes buffer, pH 7.0, containing 13 mM mercaptoethanol at 4°C. The homogenate was centrifuged at 10,000 g for 30 min at 4°C. The resulting clear supernatant was used directly for APase assay. APase activity was assayed using p-nitrophenyl phospate as substrate (Huang and Kao, 1991). One unit of APase activity is defined as the amount of enzyme which liberates 1 µmol of p-nitrophenol min-1. Pi was extracted with perchloric acid (2.5%) and determined by spectrophotometric method of Yoshida et al. (1972). All data for control, NaCl and sorbitol treatments were expressed on the basis of fresh weight, whereas those for water stress treatment were on the basis of initial fresh weight.
Native PAGE was performed using a modification of the method of Davis (1964). Final acrylamide monomer concentrations in the slab gels were 7.5% (w/v) for the separating gel and 5% (w/v) for the stacking gel. Samples were run at a constant current of 20 mA at 4°C. Following electrophoresis, APase were visualized at room temperature using 0.1% Fast red TR salt and 1% a-naphthylphosphate in 5 mM MgCl2 and 200 mM sodium acetate (pH 5.0).
Results and Discussion
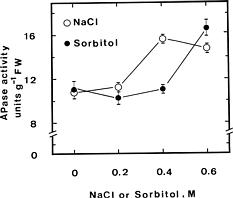
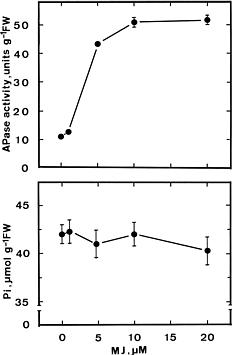
Effect of NaCl and sorbitol on APase activity in detached rice leaves under light conditions is shown in Fig
1Corresponding author. Fax: 886-2-23620879; E-mail: kaoch@cc.ntu.edu.tw