Bot. Bull. Acad. Sin. (1998) 39: 251_254
Lee and Ko Macadamia quick decline
Phytoplasmas are not associated with quick decline of
macadamia trees in Hawaii
Ing-Ming Lee1 and Wen-Hsiung Ko2,3
1Molecular Plant Pathology Laboratory, Agriculture Research Service, USDA, Beltsville, Maryland 20705, USA
2Department of Plant Pathology, Beaumont Agricultural Research Center, University of Hawaii at Manoa, Hilo, Hawaii 96720, USA
(Received January 5, 1998; Accepted April 9, 1998)
Abstract. Association of phytoplasma with quick decline of macadamia trees in Hawaii was reevaluated. Twenty samples were collected from symptomatic and symptomless trees of the two susceptible cultivars cv. 333, and cv. 344 grown at Keaau and Kaiwiki on the island of Hawaii. Ultrasensitive nested-PCR assays using two universal primer pairs failed to detect phytoplasmas in all samples tested, contradicting the recent reports that phytoplasma may play a role in macadamia decline or quick decline. In contrast, infection of trunks by two fungal pathogens, Nectria rugulosa and Xylaria arbuscula were consistently associated with declining but not healthy macadamia trees, supporting the previous suggestion that sudden death of macadamia trees is primarily caused by girdling of trunks resulting from infection by fungal pathogens.
Keywords: Macadamia integrifolia; Nectria rugulosa; Phytoplasma; Quick decline; Xylaria arbuscula.
Introduction
Quick decline of macadamia (Macadamia integrifolia Maiden & Betche) trees has become a serious problem in Hawaii in recent years. The infected trees die within 2_3 months after the appearance of initial foliage symptoms (Ko and Kunimoto, 1991a). This is different from the dieback and gradual decline of macadamia trees resulting from root rot caused by Kretzschmaria clavus (Fr.) Sacc. (Ko et al., 1977) and Ganoderma lucidum (W. Curt. ex Fr.) Karsten (Ann and Ko, 1988). Several macroscopic fungi frequently observed on trees showing quick decline symptoms were tested following Koch's rules and found to be among the causal organisms of the disease (Ko and Kunimoto, 1991a,b; 1996). Microscopic Phytophthora capsici Leonian (Ko and Kunimoto, 1994) and Acremonium recifei (Leao & Lobo) Gams (Ko and Kunimoto, 1997) frequently isolated from trunks of declining trees in certain orchards were tested with the same procedure and were also found to be the causal organisms of the disease. Recently, phytoplasmas (previously known as mycoplasmalike organisms) were reported to be associated with macadamia trees showing symptoms of quick decline (Borth et al., 1994a,b). Since symptoms of zonal lesions and girdling of trunks on declining macadamia trees (Ko and Kunimoto, 1991a,b; 1994; 1996) have never been reported to be induced by phytoplasma infection, the association of phytoplasmas with quick decline of macadamia trees was reevaluated in this study using the ultrasensitive methods for general detection of
phytoplasmas. A brief account of this work has been published (Lee and Ko, 1997).
Materials and Methods
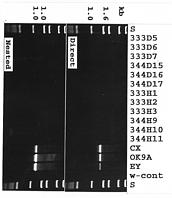
Asymptomatic (H) and symptomatic (D) (with quick decline syndrome) macadamia trees were sampled from two cultivars, cv. 333 and cv. 344 grown at Keaau and Kaiwiki on the island of Hawaii (Table 1). Direct and nested-polymerase chain reaction (PCR) assays using the two universal primer pairs, R16mF2/R16mR1 and R16F2n/R16R2, previously designed (Gundersen and Lee, 1996; Lee at al., 1993b) for general detection of phytoplasmas were employed to detect the presence of phytoplasmas in afftected leaf midribs. Total nucleic acid was extracted as previously described (Lee et al., 1993a) from apparently healthy (asymptomatic) or symptomatic macadamia leaves (midribs) collected from two cultivars on the island of Hawaii. Nucleic acid samples were diluted in sterile deionized water to give a final concentration of 20 ng/µl. Direct-PCR assays using primer pair R16mF2/R16mR1 were performed as previously described (Lee et al., 1995), with 20 ng of total nucleic acid, 200 µM each dNTP and 0.4 to 1.0 µM primer pair. Thirty-five PCR cycles were conducted in an automated thermocycler (Perkin-Elmer Cetus, Norwalk, CT). The following parameters were used: 1 min (2 min for the first cycle) denaturation step at 94°C, annealing for 2 min at 60°C (55°C for nested-PCR), and primer extension for 3 min (10 min in final cycle) at 72°C. Nucleic acid samples prepared from X-diseased phytoplasma (strain CX)- and aster yellows phytoplasma (strain OK9A)-infected periwinkle plants (Catharanthus roseus [L.] G. Don ) and elm
3Corresponding author. Tel: (808) 974-4105; Fax: (808) 974-4110.