Bot. Bull. Acad. Sin. (1999) 40: 1_7
Sisler and Serek Compounds controlling the ethylene receptor
(Invited review paper)
Compounds controlling the ethylene receptor
Edward C. Sisler1,3 and Margrethe Serek2
1Department of Biochemistry, North Carolina State University, NCSU Box 7622, Raleigh, NC 27595-7622, USA
2Department of Agricultural Sciences, Horticulture, Royal Veterinary and Agriculture University, Thorvaldsensvej 57, DK-1871 Frederiksberg C., Denmark
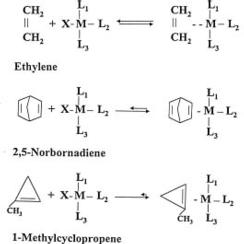
Abstract. Many organic compounds interact with the ethylene receptor and diffuse free after different periods of time. Some are agonists and mimic ethylene, and some are antagonists, which prevent ethylene action by blocking the receptor. Some of the antagonists have proven useful in scientific studies, and some promise to be commercially important in protecting against ethylene. The times that different compounds remain bound may be important clues to how ethylene transmits its signal to the signal transduction pathway. Ethylene diffuses from the site with a t1/2 of 2_10 minutes and is an active compound. For 2,5-Norbornadiene (2,5-NBD) and some other strained olefins that block ethylene action, the half diffusion time is 3_6 h. For other more strained compounds (cyclopropenes) the half diffusion time is estimated to be 7_12 days; they block ethylene action during this time. The time of diffusion from the receptor appears to be the major difference between compounds that block the receptor and those that are active. It is suggested that this time constant may be the controlling factor in ethylene action.
Keywords: 1-methylcyclopropene, 1-MCP; 2,5-norbornadiene, 2,5-NBD; 3,3-dimethylcyclopropene, 3,3-DMCP; Cyclopropene; Diazocyclopentadiene, DACP; Ethylene antagonist; Ethylene receptor; Methylenecyclopropane; trans-Cyclooctene.
Abbreviations: 1-MCP, 1-methylcyclopropene; 2,5-NBD, 2,5-norbornadiene; 3,3-DMCP, 3,3-dimethylcyclopropene; DACP, diazocyclopentadiene.
Contents
Introduction 1
Uses of Commercial or Scientific Value 2
Effects of Concentration 3
Competition for the Receptor 4
How Long do Compounds Remain Bound to the Receptor? 4
Why do Olefins Bind to Metals? 4
How do these Compounds Prevent Ethylene Action? 5
What are the Ligands? 5
How does the Active Complex Work? 5
Where is the Site of Ethylene Action? 5
Future Work 6
Literature Cited 6
Introduction
In recent years some very effective agents for blocking the ethylene receptor have been discovered by Sisler and coworkers. These block ethylene action, and hold the promise of being a new way of controlling ripening, se
nescence, and other ethylene responses. Since the ethylene receptor is ubiquitous in plants, these compounds should control all ethylene responses in plants. Rather than being a totally new discovery, they are the consequence of efforts to develop a better understanding of the ethylene receptor. This paper is not intended to cover all of the many papers in which these compounds have been used, but instead to cover the important findings and uses of the compounds. We will initially address what has been
3Corresponding author. Tel: (919) 515-5761; Fax: (919) 515-2047; E-mail: sisler@bchserver.bch.ncsu.edu