Bot. Bull. Acad. Sin. (1999) 40: 87_92
Carballeira and Reigosa Allelopathic natural leachates of Acacia dealbata
Effects of natural leachates of Acacia dealbata Link in Galicia (NW Spain)
Alejo Carballeira1 and Manuel J. Reigosa2,3
1Ecología, Departamento de Biología Fundamental, Facultad de Biología, Universidade de Santiago de Compostela, Spain
2Fisioloxía Vexetal, Dep. Bioloxía Vexetal e Ciencia do Solo, Facultade de Ciencias de Vigo, Universidade de Vigo, Apt 874, 36200 Vigo, Spain
(Received October 1, 1997; Accepted July 2, 1998)
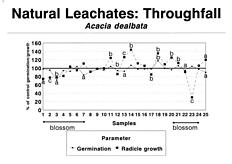
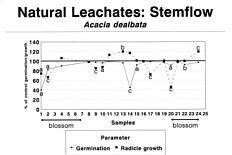
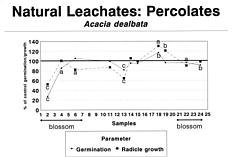
Abstract. A systematic collection of throughfall (rain passing through the canopies), stemflow (rain flowing over the stems), and soil percolates was carried out in a Acacia dealbata plantation during one year, and its allelopathic capacity was biotested using Lactuca sativa L. var. Great Lakes. The results showed an allelopathic effect related to flowers: Germination of L. sativa was inhibited 30% by throughfall, 60% by stemflow, and 75% by soil percolates during the blossoming of A. dealbata, while radicle growth of L. sativa was inhibited 23%, 33%, and 48% by the same solutions. Climatic factors, particularly rainfall some days before, were also important. The three types of samples showed significant toxicities during some periods of the year, particularly percolates, which retained toxicity for a longer time, due perhaps to the decomposition of flowers fallen on the ground.
Keywords: Acacia dealbata; Allelopathy; Percolate; Stemflow; Throughfall.
Introduction
Certain allelopathic phenomena result when toxins leach into rainfall or dew (Carral et al., 1988; Del Moral and Muller, 1969; Juste et al., 1985; Molina et al., 1991; Rama Devi et al., 1997). Demonstration of allelopathy generally has been accomplished through biotests "made in laboratory" (Hibbs and Schumaker, 1987; Leather and Einhellig, 1987; Rice, 1984). The validity of some of these tests is debatable due to the artificial manner in which the solutions of the supposed allelopathic agents were obtained (Harper, 1977; Inderjit and Dakshini, 1995; Stowe and Kil, 1981; Wardle, 1987). It is often difficult in some bioassays reported in the literature to know if the concentrations of allelochemicals tested are ecologically meaningful. There is often a lack of correlation between artificial assays and field experimental data (Henn et al., 1988; Harper, 1977; Stowe, 1979).
Some species of the genus Acacia have shown phytotoxic capacity (Duham and Lakshminarayana, 1995; Elakovich and Wooten, 1995; González et al., 1995; Nsolomo et al., 1995; Puri et al., 1994; Saxena and Sharma, 1996; Souto et al., 1994, 1995).
Acacia dealbata Link is a leguminous plant coming from Australia and Tasmania. Introduced in the European continent in 1820, it is very common in Mediterranean countries where it is both found in the wild and farmed for ornamental use. Acacia dealbata can become a blight
(García, 1979). It invades vineyards, grows in meadows, destroys retaining walls, pulls archaeological excavations down and makes access to natural areas difficult. In addition to its great colonizing capacity it leads to a very low covering and scarcity of undergrowth species. Based on this, the existence of allelopathy was first suspected. A priori, the probability of such hypothesis was supported by the fact that, as Rabotnov (1974) said, "allelopathic phenomena are chiefly possible between species originating in different regions."
A later series of experiments (Casal et al., 1985; Reigosa et al., 1984) showed A. dealbata to have a great potential allelopathic capacity through several mechanisms fluctuating within the year and within short periods of time. These fluctuations could have important ecological consequences if the highest moments of toxin production coincided with the most critical period for the receiver (Muller, 1970).
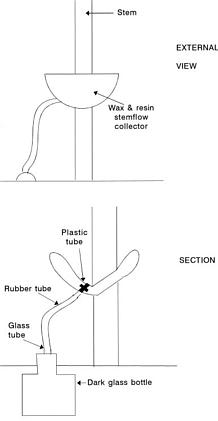
Rice (1984) indicated that it is necessary to get more information concerning rates of production and escape to the environment of the allelopathic compounds. In this paper we report that during one year we collected the natural throughfall (water that passes through the canopy and falls to the soil), stemflow (water fraction that arrives to the soil flowing over the stems), and percolates, recording data on rain episodes and nutrient leakage. Our objective was to test the possibility that allelopathic phenomena occurred in response to toxins and also to determine the influence of the phenological phase of the A. dealbata and meteorological factors on the allelochemical effects.
3Corresponding author. Tel: +34-86-812594; Fax: +34-86-812556; E-mail: mreigosa@uvigo.es