Bot. Bull. Acad. Sin. (1999) 40: 101_106
Huang et al. Infection of pollen by Botrytis cinerea
Infection of alfalfa pollen by Botrytis cinerea
H.C. Huang1, E.G. Kokko, and R.S. Erickson
Research Centre, Agriculture and Agri-Food Canada, P. O. Box 3000, Lethbridge, Alberta, Canada T1J 4B1
(Received May 6, 1998; Accepted July 24, 1998)
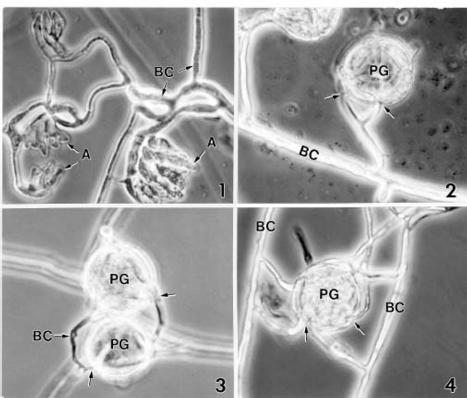
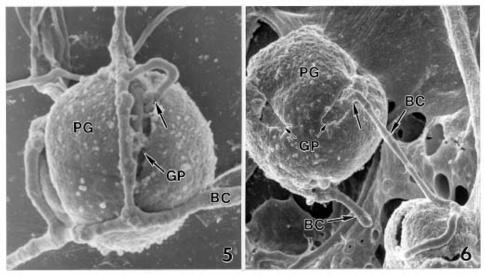
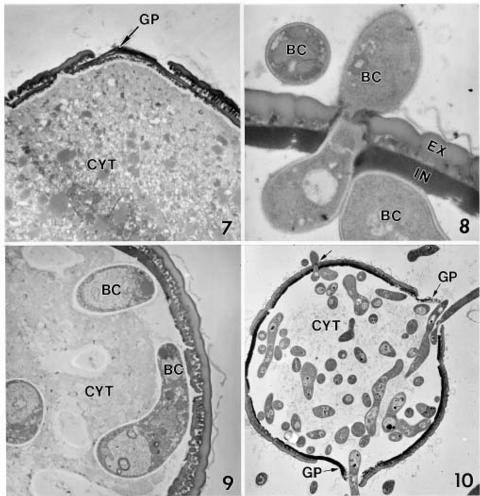
Abstract. A microscopic study proved that pollen grains of alfalfa are susceptible to infection by Botrytis cinerea, an important pathogen for blossom blight of alfalfa in western Canada. Pollen grains were inoculated by mixing pollen with drops of a B. cinerea spore suspension on glass slides. Infection occurred by direct hyphal penetration of the pollen cell walls without the formation of appressoria. Infection through the germ pores was observed more frequently than through other parts of the pollen walls. Plasmolysis of the plasma membrane and destruction of the cytoplasm occurred as a result of intensive growth of hyphae in the cell lumen of infected pollen grains. The impact of pollen infection by B. cinerea on the epidemiology of blossom blight of alfalfa is discussed.
Keywords: Alfalfa; Blossom blight; Botrytis cinerea; Epidemiology; Infection of pollen.
Introduction
Botrytis cinerea Pers.:Fr. is an important pathogen causing blossom blight, pod rots, and fruit rots on many crops, including alfalfa (Medicago sativa L.) (Gossen et al., 1996; 1997). Numerous reports indicate that high disease incidence and severity are associated with the colonization of senescent petals by B. cinerea (Bachelder and Orton, 1962; Hartill and Campbell, 1974; Hunter et al., 1972; Jarvis and Borecka, 1968; Kunimoto et al., 1996; Ogawa and English, 1960). Some reports indicate that pollen stimulates spore germination (Hartill, 1975) and infection by B. cinerea (Bachelder and Orton, 1962; Chou and Preece, 1968; Ogawa and English, 1960).
Recent studies have shown that alfalfa pollen is susceptible to infection by plant pathogens such as Verticillium albo-atrum Reinke & Berthold (Huang and Kokko, 1985) and Sclerotinia sclerotiorum (Lib.) de Bary (Huang et al., 1997). The objective of this study was to determine the susceptibility of alfalfa pollen to B. cinerea, and to describe the mode of infection using light microscopy (LM) and electron microscopy.
Materials and Methods
Inoculation of Alfalfa Pollen
Seeds of alfalfa, cultivar Barrier, were sown in Cornell peat-lite mix (Boodley and Sheldrake, 1977) in plastic pots (15 cm diam.), and plants were grown to maturity in a greenhouse. Alfalfa flowers were excised and tripped to collect fresh pollen grains on glass slides. An aqueous spore suspension containing approximately 103 spores/ml
of B. cinerea was made from 3-week-old cultures grown on potato dextrose agar in petri dishes. Pollen grains were inoculated with B. cinerea by mixing the pollen with five drops of the suspension on each slide. The slides were placed in a moist chamber and kept for five days at room temperature (20_22°C) before being prepared for microscopic examinations. Two controls, pollen grains in sterile water and B. cinerea spores in sterile water, were prepared and processed for microscopic examinations.
Light Microscopy
For light microscopy, each 5-day-old sample (slide) of pollen-spore mixture was mounted in lactophenol and covered with a cover slip. The mounted specimens were examined for infection of pollen by B. cinerea using a Zeiss Photomicroscope III. Control samples were also mounted and examined microscopically. The important features observed in the specimens were documented by phase contrast photomicrographs.
Scanning Electron Microscopy (SEM)
For SEM, the 5-day-old inoculated (B. cinerea) and uninoculated (control) pollen samples were immersed in 2% glutaraldehyde fixative in 0.05 M sodium phosphate buffer, pH 7.0, at 4°C overnight (16 h) and then brought to room temperature. The samples were washed (3 × 10 min) with the sodium phosphate buffer solution. The specimens were dehydrated in a graded series of ethanol and critical point dried (Polaron E3100) with liquid carbon dioxide as the transitional fluid. The material was adhered onto aluminum specimen mounts with colloidal silver paste, air-dried overnight and sputter-coated (Denton Vacuum Desk-1) with gold (approximately 15 nm thickness). The specimens were examined and photographed on a Hitachi S-570 SEM.
1Corresponding author. Fax: (403) 382-3156; E-mail: huangh@ em.agr.ca