Bot. Bull. Acad. Sin. (1999) 40: 121-126
Sammour Proteins of linseed
Proteins of linseed (Linum usitatissimum L.), extraction and characterization by electrophoresis
Reda Helmy Sammour1
Department of Botany, Faculty of Science, Tanta University, Tanta, Egypt
(Received March 17, 1998; Accepted August 13, 1998)
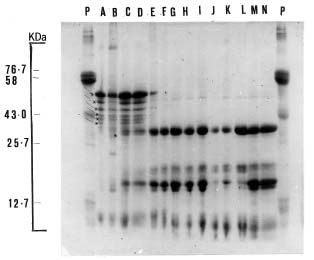
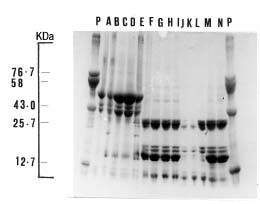
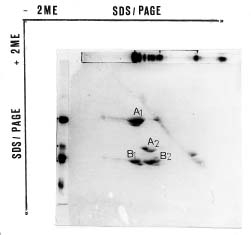
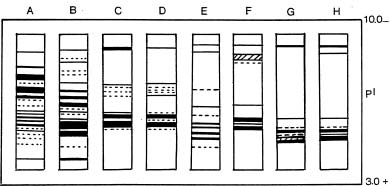
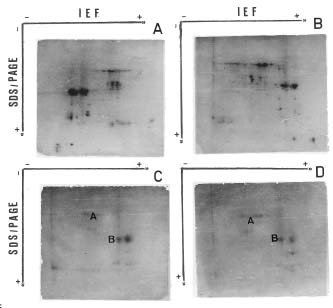
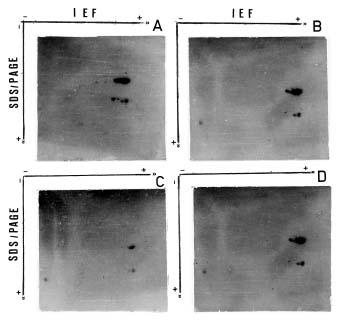
Abstract. The seed proteins of linseed (Linum usitatissimum L.) were qualitatively and quantitatively analyzed. Qualitative studies were carried out using an array of electrophoretic techniques, including sodium dodecylsulphate polyacrylamide gel electrophoresis (SDS-PAGE), sodium dodecylsulphate porosity gradient polyacrylamide gel electrophoresis (SDS-Poro-PAGE), two dimension sodium dodecylsulphate polyacrylamide gel electrophoresis (2-D SDS-PAGE), isoelectric focusing, and mapping gels. Electrophoretic patterns of the proteins extracted with water, buffer, urea, and SDS, and analyzed on SDS-PAGE and Poro-SDS-PAGE under non-reducing conditions, showed six major bands with MWs of 55 kDa, 50 kDa, 47 kDa, 45 kDa, 43 kDa and 41 kDa. After reduction with 2-mercaptoethanol (2-ME), these bands gave rise to large acidic chains with MWs around 40 kDa and small basic chains with MWs around 20 kDa. The pI-values of linseed proteins are distributed over the pH range used, namely pH 3_10. Mapping gels showed that the major bands were highly heterogeneous. Quantitative estimation of the different protein species in the seed flour indicated that the contents of albumin, globulin, prolamin, and glutelin were 197 ± 19, 196 ± 12, 32 ± 7, and 65 ± 6 mg/g seed flour, respectively.
Keywords: Albumin; Globulin; Kilodalton (kDa); 2-mercaptoethanol (2-ME); Molecular weights (MWs); Two dimension-PAGE (2-D SDS-PAGE).
Abbreviations: SDS-PAGE, sodium dodecylsulphate polyacrylamide gel electrophoresis; SDS-Poro-PAGE, sodium dodecylsulphate porosity gradient polyacrylamide gel electrophoresis; 2-D SDS-PAGE, two dimension sodium dodecylsulphate polyacrylamide gel electrophoresis; 2-ME, 2-mercaptoethanol; PAGIF, polyacrylamide gel isoelectric focusing.
Introduction
Linseed or flax (Linum usitatissimum L.) is an annual crop belonging to the family Linaceae (Simmonds, 1976). It is grown worldwide either for the oil extracted from the seed or for fiber from the stem. The meal that remains after oil is extracted from the seed is fed to animals as a protein supplement (Lennerts, 1983). Linseed meal is 35_40% protein and together with cottonseed and sunflower supplies about 23% of the world's oilcake/meal (Hatje, 1989).
Lay and Dybing (1989) improved seed yield and oil production through breeding procedures that aimed to improve disease resistance, notably against rust and wilt. However, the agronomically important traits, such as seed yield and its components, and oil yield, have received little research effort.
The seed protein of flax was purified by ultracentrifugation (Youle and Huang, 1981). It was found to contain two kinds of proteins; 2S protein (albumin) and 11S (globulin) protein (the major storage protein). However, few qualitative studies have emphasized the components of linseed proteins. This work, therefore, is devoted to bridging this gap in the literature.
Materials and Methods
Materials
Linseed (Linum usitatissimum L. cultivar Viking) was obtained from Cooperative Liniere De Fontaine-Cany, France. The seeds were ground with an electric mill (JANKE and KUNKEL, type) for 1 min at 0°C. The flour was defatted by stirring with pure hexane (1:10 W/V) for 30 min three times. After filtering, the flour was air-dried, brushed through a sieve of 125 µm (115 mesh), and stored at -10°C prior to use.
Methods
Seed flour extracts
Water, buffer, SDS, and urea extracts were made under non-reducing and reducing conditions (with 1% 2-ME). To made extracts under either reducing condition, 2-ME was added to a final concentration of 1% at the onset of extraction.
Water and buffer extracts. Portions (60 mg) of air dried linseed flour were extracted overnight with 500 µl distilled water and 0.125 M Tris/borate buffer pH 8.9 (containing 0.02% sodium azide) in an Eppendorf tube at room temperature. The mixture was centrifuged for 20 min at 4°C and 10,000 g (Heraeus Christ Labofuge I-cooling centrifuge). A second water extract was made,
1Corresponding author: Teacher's College, Sakaka El-Jouf, P.O. Box 957, Saudi Arabia. Tel: (04) 6250471.