Bot. Bull. Acad. Sin. (1999) 40: 159-164
Hur et al. Genetic diversity of G. japonica
Genetic diversity and population structure of Gleditsia japonica var. koraiensis in Korea
Man Kyu Huh1,3, Hak Young Lee2, and Hong Wook Huh1
1Department of Biology Education, Pusan National University, Pusan, 609-735, The Republic of Korea
2Department of Biology, Chonnam National University, Kwangju 500-757, The Republic of Korea
(Received June 29, 1998; Accepted September 23, 1998)
Abstract. Enzyme electrophoresis was used to estimate genetic diversity and population genetic structure of Gleditsia japonica var. koraiensis in Korea. Eleven of the 17 loci (64.7%) showed detectable polymorphism. Genetic diversity (0.247) was higher than average for species with similar life history traits. Analysis of fixation indices, calculated for all polymorphic loci in each population, showed a substantial deficit of heterozygotes relative to Hardy-Weinberg expectations. This deficit is partly associated with inbreeding which is due to consanguineous and selfing mating. The average GST for polymorphic loci was 0.145, indicating that most (85.5%) of the genetic diversity occurred within populations. The indirect estimate of gene flow based on mean GST was moderate (Nm = 1.24). Given limited gene flow, populations are expected to diverge genetically due to drift, the random loss of alleles due to small population size.
Keywords: Genetic diversity; Gleditsia japonica; Population structure.
Introduction
During the past 20 years, enzyme electrophoresis has been used to describe the population genetic structure of over 700 plant taxa (Hamrick and Godt, 1989). This information has contributed greatly to an understanding of the evolutionary history of individual species and related group of species (Haufler, 1987) and has provided insights into the relationships between allozyme diversity and life-history traits (Loveless and Hamrick, 1984; Kiang and Chiang, 1990). Recently, allozyme studies of woody angiosperms have found that these plants are comparable to conifers with respect to their ecological and/or life history trait (e.g. Schnabel and Hamrick, 1990a,b; Sherman-Broyles et al., 1992). These studies have shown that such angiosperms have high levels of genetic variation, and a low proportion of their genetic diversity is among populations. Generalizations derived from the allozyme literature provide a basis on which to build sound programs for the conservation of genetic diversity of rare and endangered species (Hamrick et al., 1991). In addition, allozyme diversity can be used as a yardstick to measure the effectiveness of in situ and ex situ conservation programs (Hamrick et al., 1991). Despite the importance of knowledge on genetic variation for providing information for conservation purposes, detailed studies of genetic variation are not available for most native taxa in China and Korea, particularly woody plants (Kang and Chung, 1997). In addition, almost no information is available from flora-rich countries in Africa or from China (Bennett and Leich, 1995).
Gleditsia japonica Miquel var. koraiences Nakai is a polygamous woody plant that is distributed in natural habitats of mountains. It has a strong spike in stem and branches, and polygamous flowers are present in separate inflorescences. Gleditsia japonica var. koraiences is not an abundant plant over its range in Korea (Hur and Lee, personal observation). Furthermore, young plants are rare in most populations. Population size is highly variable, ranging from a few individuals at the margins of forests to several dozen plants in valleys on high-elevated mountains. Populations that are reproductively isolated may gradually exhibit genetic differentiation. The rapid loss of new plants results in the permanent loss of gene pools with potential applications in medicine and species conservation. Gleditsia japonica var. koraiensis is an economically useful species in China and Korea, as the seeds have been used in medicine. The purposes of this paper are: 1) to estimate how much total genetic diversity is maintained in the species, 2) to describe how genetic variation is distributed within and among populations, and 3) to compare our estimates with those for species having very similar life history traits.
Materials and Methods
Sampling Procedure and Enzyme Electrophoresis
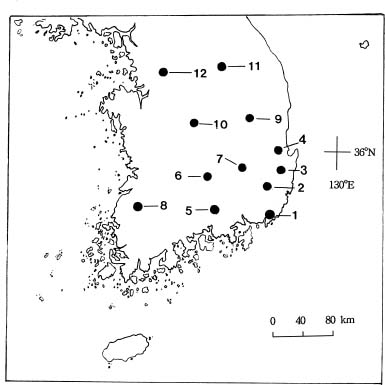
Seeds of Gleditsia japonica Miquel var. koraiensis Nakai were collected from twelve natural populations in Korea between 1995 and 1996 (Figure 1). Twenty to thirty legumes were collected from each population, and one seed from each legume was used in this study. Seeds were moistened with 98% sulphuric acid for 30 min, rinsed with tap water, and incubated at 20°C for germination. Ger
3Corresponding author. Tel: 51-510-2698; Fax: 82-51-514-8576.