Lin and Kao Copper and putrescine accumulation
Bot. Bull. Acad. Sin. (1999) 40: 213-218
Excess copper induces an accumulation of putrescine in rice leaves
Chuan Chi Lin and Ching Huei Kao1
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
(Received September 2, 1998; Accepted January 15, 1999)
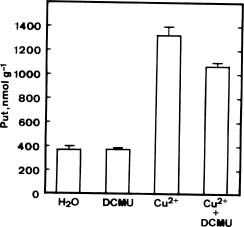
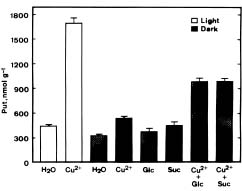
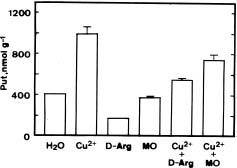
Abstract. The effect of excess Cu2+ (sulfate salt) on putrescine (Put) accumulation in detached rice leaves was investigated. Cu2+ treatment increased Put concentrations in rice leaves under both light and dark conditions. This increase was more pronounced in the light than in the dark, suggesting the importance of illumination in Cu2+-induced accumulation of Put in detached rice leaves. The photosynthetic electron transport inhibitor 3-(3,4-dichlorophenyl)-1,1-dimethylurea reduced Put accumulation induced by Cu2+ in the light. In darkness, Cu2+-induced Put accumulation was more effective in the presence of glucose or sucrose than in their absence. In the light, Cu2+ also induced Put accumulation in six other rice cultivars. D-Arginine and a-methylornithine decreased concentrations of Put induced by Cu2+, indicating that Cu2+ may affect the activities of both arginine decarboxylase and ornithine decarboxylase, enzymes responsible for the biosynthesis of Put.
Keywords: Copper; Putrescine; Oryza sativa.
Abbreviations: D-Arg, D-arginine; DCMU, 3-(3,4-dichlorophenyl)-1,1-dimethylurea; MO, a-methylornithine; Put, putrescine; Spd, spermidine; Spm, spermine.
Introduction
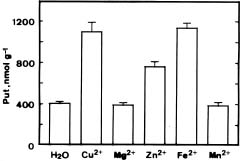
Polyamines are low-molecular weight polycations present in all living organisms. Putrescine (Put) is the obligate precursor for spermidine (Spd) and spermine (Spm) in all systems studied so far (Evans and Malmberg, 1989). In response to various types of environmental stress, plants accumulate Put (Evans and Malmberg, 1989). In recent decades, industrial and urban activities have increased the deposition of heavy metals (such as copper) in the soil system (Tyler, 1972). Despite the apparent importance of polyamines, especially Put, in stress metabolism, little information is available on the effects of heavy metals on plant polyamine contents. Cd2+ treatment of detached oat and rice leaves resulted in a significant increase in Put concentrations (Hou and Kao, 1993; Weinstein et al., 1986). Agrawal et al. (1992) demonstrated that exposure of an unicellular green alga to mercury caused an increase in Put concentrations.
Copper is an essential element for plant growth (Arnon and Stout, 1939) and important in various biochemical process, but at toxic concentrations it interferes with numerous physiological processes (Fernandes and Henriques, 1991). Virtually no information has been reported on the effect of Cu2+ on the accumulation of polyamines. The objective of this study was to examine the effect of Cu2+ on polyamine concentrations in detached rice leaves.
Materials and Methods
Rice (Oryza sativa L. cv. Taichung Native 1) was cultured as previously described (Kao, 1980). The apical 3-cm segments excised from the third leaves of 12-day-old seedlings were used. A group of 10 segments was floated in a Petri dish containing 10 mL deionized water or aqueous solution of tested compounds. Detached rice leaves were treated with 0.01~10 mM CuSO4 at 27°C under light (40 µmol m-2 s-1). For other experiments, incubation was carried out for various lengths of time in the light or in the dark.
For polyamine extraction, leaf segments were homogenized in 5% (v/v) perchloric acid. Polyamines were determined using HPLC after benzoylation as described previously (Chen and Kao, 1991). The amounts of polyamines were expressed as nmol g-1 fresh weight.
All experiments were performed three times and within each experiment treatments were replicated 4 times. Similar results and identical trends were obtained each time. The data reported here are from a single experiment.
Results
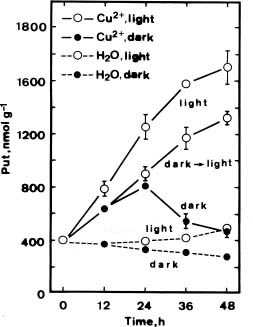
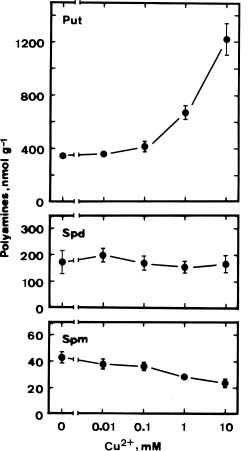
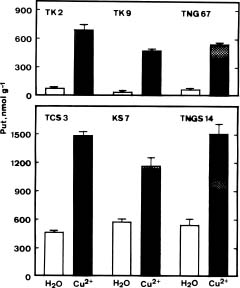
As indicated in Figure 1, the amount of Put increased significantly at concentrations of 1 and 10 mM Cu2+. After 36 h incubation under light, treatment with Cu2+ at 10 mM, increased Put concentration 3- to 4-fold. No significant changes in Spd concentrations were observed in Cu2+-treated detached rice leaves, whereas the concentrations of Spm were almost halved at the 1 and 10 mM Cu2+.
1Corresponding author.