Bot. Bull. Acad. Sin. (1999) 40: 227-230
Chung Allozyme variation in Lycoris radiata
Notes on allozyme variation in Lycoris radiata (Amaryllidaceae) from Korea
Myong Gi Chung
Department of Biology, Gyeongsang National University, Chinju 660-701, The Republic of Korea
(Received July 23, 1998; Accepted January 5, 1999)
Abstract. Lycoris radiata (L'Her) Herb. var. radiata, a herbaceous perennial and sterile triploid (2n = 33), grows in moist places in Japan (except Hokkaido), Korea, China, and Nepal. Several botanists have speculated that the taxon originated in China and was introduced into Japan. In Korea, the taxon is routinely found in the streamsides near temples in the southern regions. Allozyme analysis was conducted in eight Korean populations of L. radiata var. radiata as well as in one Korean population of L. chinensis Traub. (2n =16) to estimate levels of allozyme diversity. Lycoris radiata var. radiata was monomorphic at all 24 allozyme loci surveyed whereas L. chinesis was polymorphic at 19% of the 21 loci analyzed with 0.06 gene diversity. The results strongly indicated that only one or a few bulbs of L. radiata var. radiata were introduced from China or, secondarily, Japan to a temple in Korea and were then naturalized in the southern Korean peninsula via a strong vegetative reproduction by the rapid formation of new bulbs.
Keywords: Allozyme; Korea; Lycoris chinensis; L. radiata var. radiata; Naturalization; Sterile triploid.
Introduction
Electrophoretic techniques provide genetic markers to recognize individual genotypes for asexually reproducing plants (e.g., Pleasants and Wendel, 1989; Murawski and Hamrick, 1990; Aspinwall and Christian, 1992; Lokker et al., 1994; Kim and Chung, 1995). This approach has made it possible to better understand the spatial distributions of clones and the genotypic diversity maintained within populations (e.g., Parker and Hamrick, 1992; Berg and Hamrick, 1994). Allozyme analysis has contributed in a variety of ways to better understanding of the origin, history of naturalization or domestication, and speciation of plant species (Crawford, 1989; Doebley, 1989).
Lycoris radiata (L'Her.) Herb. (Amaryllidaceae) consists of three varieties: L. radiata var. radiata, L. radiata var. pumila Grey, L. radiata var. kazukoana Yonezawa (Hsu et al., 1994). The former variety is the most widespread taxon of Lycoris. It is distributed widely in China, Japan (except Hokkaido), and the southern Korean peninsula. Lycoris radiata var. pumila, a fertile diploid (2n=22), is endemic to eastern, central, and southern China (Hsu et al., 1994). Lycoris radiata var. kazukoana is endemic to Japan (Yonezawa, 1989).
Lycoris radiata var. radiata is a sterile triploid (2n=33) that propagates via strong vegetative reproduction by the rapid formation of new bulbs. It has been suggested, based on karyotype analysis, that this variety might have originated from the combination of an unreduced gamete
of a diploid with a normal gamete of a diploid L. radiata var. pumila (Liu and Hsu, 1989) because tetraploids have never been found in this species. It has also been suggested that L. radiata var. radiata originated in China and was introduced into Japan (Fukuda et al., 1980; Kurita, 1987) because the diploid L. radiata var. pumila has never been found in Japan or its adjacent regions (Hsu et al., 1994). Lycoris radiata var. radiata is routinely found in the streamsides near temples in southern Korea (Park et al., 1986b; M. Chung, pers. obs.).
In this study, allozyme analysis was conducted on eight Korean populations of L. radiata var. radiata as well as on one population from one known locality of L. chinensis Traub. (2n =16) in Korea to estimate levels of allozyme and clonal diversity and to gain ideas concerning the origin of this variety in Korea.
Materials and Methods
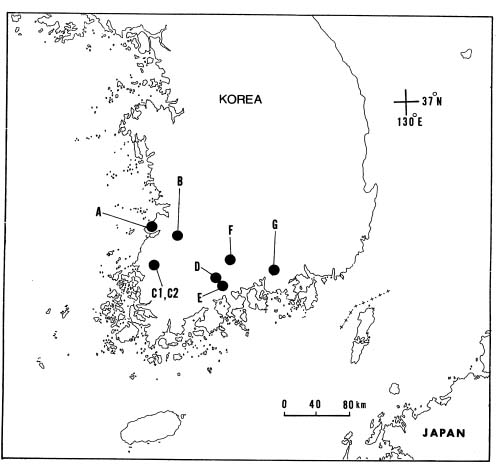
Leaves were collected from eight populations of L. radiata var. radiata in Korea (Figure 1). Fifty individuals per population were randomly sampled in a 1,000~2,000 m2 area, depending on population size. Because the variety exhibits extensive clonal growth, samples were collected at intervals of > 3 m within each population to avoid biasing samples toward certain clones. In addition, twenty-two individuals of L. chinenis were collected from hillsides of Mt. Dosol, Kochang Gun, Pref. Chollabuk, Korea. Leaf samples were placed in plastic bags wrapped with a wet paper towel and stored on ice and transported to the laboratory. Samples were then stored at 4°C until protein extraction.
Fax: +82-591-54-0086; E-mail: mgchung@nongae.gsnu.ac.kr