Bot. Bull. Acad. Sin. (1999) 40: 289-294
Chang and Lee Proline in Gracilaria tenuistipitata
High temperature-induced free proline accumulation in Gracilaria tenuistipitata (Rhodophyta)
Yuan-Chun Chang and Tse-Min Lee1
Institute of Marine Biology, National Sun Yat-sen University, Kaohsiung 80424, Taiwan
(Received November 23, 1998; Accepted February 12, 1999)
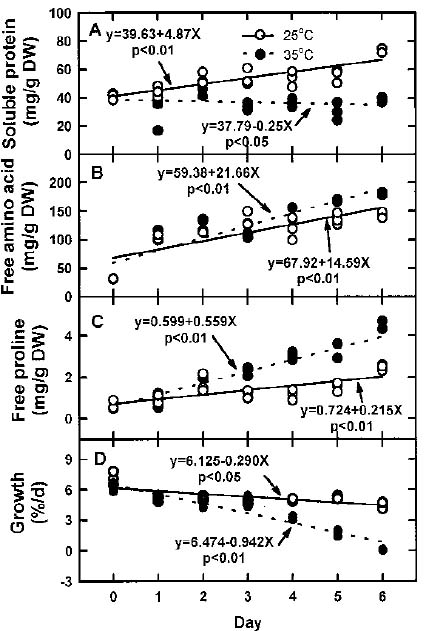
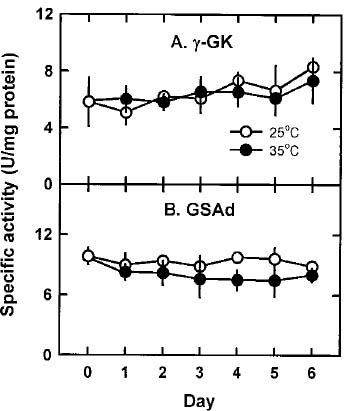
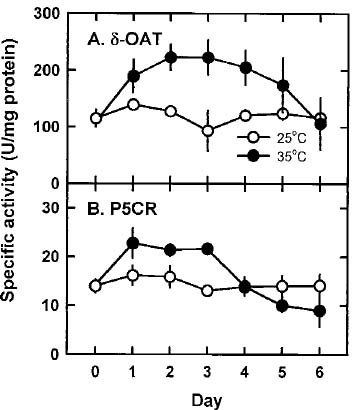
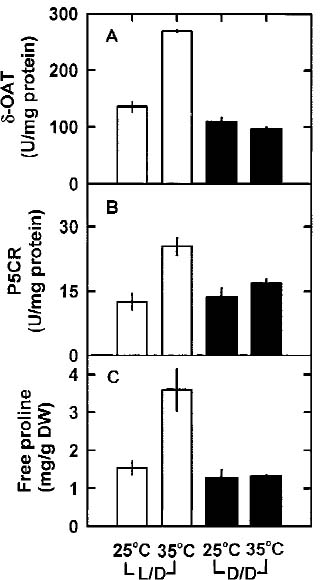
Abstract. The effects of high temperature (35°C) on free proline biosynthesis were studied in a marine red macroalga Gracilaria tenuistipitata var. liui Zhang et Xia. Exposure to 35°C decreased the specific growth rate after 3 days and increased the free proline levels after 2 days, but did not affect the free amino acid levels. Soluble protein levels were constant on exposure to 35°C while those of 25°C-treated thalli increased steadily through the treatment periods. The specific activities of both g-glutamyl kinase (g-GK; EC 2.7.2.11) and glutamate-5-semialdehyde dehydrogenase (GSAd; EC 1.4.1.3) remained unchanged after exposure to 35°C. Upon exposure to 35°C, the specific activity of ornithine d-aminotransferase (d-OAT; EC 2.6.11.3) increased rapidly after 2 days and decreased at Day 5, while that of D1-pyrroline-5-carboxylate reductase (P5CR; EC 1.5.1.2) also increased after 1 day and decreased at Day 4. The increase in d-OAT and P5CR specific activities coincides with the increase in free proline levels. The 35°C induction of free proline accumulation and d-OAT and P5CR activity increase is a light-dependent process. These results suggest that the stimulation of both synthesis (d-OAT and P5CR) and protein proteolysis contributes to the 35°C-induced free proline accumulation in G. tenuistipitata.
Keywords: Gracilaria tenuistipitata; GSAd; High temperature; P5CR; Proline; Protein proteolysis; Red alga; d-OAT; g-GK.
Abbreviations: g-GK, g-glutamyl kinase; GSAd, glutamate-5-semialdehyde dehydrogenase; d-OAT, ornithine d-aminotransferase; P5CR, D1-pyrroline-5-carboxylate reductase.
Introduction
In higher plants, proline accumulates under stress and shows an association with stress adaptation (Aspinall and Paleg, 1981; Lalk and Dorffling, 1985). Proline also accumulates in algae under stress (Bartels and Nelson, 1994; Kalinkina and Naumova, 1992; Singh et al., 1996; Wu et al., 1995). It has been shown in higher plants that proline is synthesized from either the glutamate pathway or the ornithine pathway (Delauney and Verma, 1993). The glutamate pathway is initiated by phosphorylating L-glutamate to L-glutamyl-g-phosphate by g-glutamyl kinase (g-GK; EC 2.7.2.11). Then, g-GK is converted to glutamate- g -semialdehyde by GSA dehydrogenase (GSAd; EC 1.4.1.3). Finally GSA spontaneously forms D1-pyrroline-5-carboxylate (P5C), and P5C is reduced to proline by P5C reductase (P5CR; EC 1.5.1.2). In the ornithine pathway, ornithine is converted to GSA by ornithine d-aminotransferase (d-OAT; EC 2.6.11.3) and then follows the same route to form P5C as in the glutamate pathway.
A stimulation of synthesis is a factor contributing to the stress-induced proline accumulation, but which pathway becomes responsible for proline synthesis is depen
dent on the plant systems involved or the kind of stress (Delauney and Verma, 1993). Under osmotic stress, the glutamate pathway is generally considered the primary route for proline synthesis (Delauney et al., 1993). However, in the case of the NaCl-tolerant Brassica juncea L., the NaCl-induced proline synthesis is suggested to be derived from the ornithine pathway with d-OAT as the principle enzyme (Madan et al., 1995). In addition, the excess nitrogen supply switches the synthesis of proline from the glutamate to the ornithine pathway (Delauney et al., 1993).
The economically important marine red alga Gracilaria tenuistipitata var. liui Zhang et Xia, which is farmed in brackish water ponds as a main food source for the cultivation of sea abalone Haliotis diversiolor supertexta Lischke in Taiwan, frequently suffers from high water temperatures in summer (Chiang, 1981). It was recently found that proline significantly accumulated in G. tenuistipitata at 35°C, its upper survival limit (Lee et al., 1999). This study attempted to investigate whether synthesis was stimulated by the 35°C-induced free proline accumulation in G. tenuistipitata. The time course of changes in the levels of soluble protein, free amino acids, and free proline were compared to discover whether protein proteolysis is involved in the free proline accumulation. The relationships between proline accumulation and specific growth rate were also compared to understand their possible roles in G. tenuistipitata exposed to high temperature. The time-
1Corresponding author. Tel: 886-7-5252000 ext. 5110; Fax: 886-7-5255100; E-mail: tmlee@mail.nsysu.edu.tw