Bot. Bull. Acad. Sin. (2000) 41: 1-10
Cheng and Yeh In vivo expression of PRSV CP genes with different leaders
Construction and evaluation of transgenic tobacco plants
expressing the coat protein gene of papaya ringspot virus
with different translation leaders
Ying-Huey Cheng1 and Shyi-Dong Yeh2,3
1Department of Horticulture, Taiwan Agricultural Research Institute, Wufeng, Taiwan, Republic of China
2Department of Plant Pathology, National Chung Hsing University, Taichung, Taiwan, Republic of China
(Received August 19, 1998; Accepted June 5, 1999)
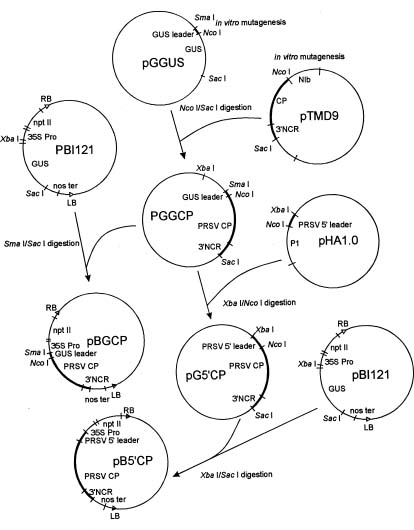
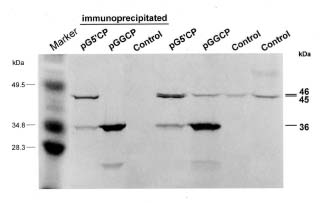
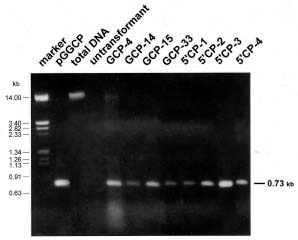
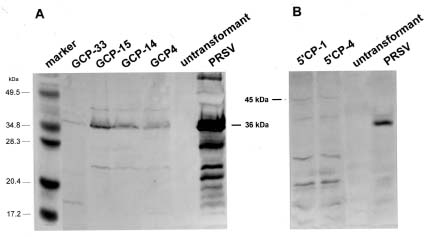
Abstract. Papaya ringspot virus (PRSV) YK isolate used in this study is a local mosaic strain isolated from Yung-Kang, Tainan, and its genome has been cloned and completely sequenced. A NcoI site before the coat protein (CP) reading frame of PRSV YK was generated by oligonucleotide-directed mutagenesis, and then the CP reading frame with the 3' noncoding region of PRSV YK was ligated with the gus leader sequence from the pGEM vector to create the construct pGGCP. To express the CP with a homologous viral translation sequence, the gus leader was replaced by the cDNA sequence corresponding to the 5' region (nt 1-347) of PRSV genome to generate a protein containing 9 kDa polypeptide of PRSV P1 protein fused with the CP, and the construct was designated as pG5'CP. In vitro translation from the transcripts derived from pGGCP and pG5'CP generated protein products of 36 kDa and 45 kDa, respectively. Both proteins reacted with the antiserum to PRSV CP, and the level of 36 kDa protein was higher than that of 45 kDa protein. The CP reading frame with the gus or PRSV 5' leaders was individually subcloned into a Ti binary vector. Transgenic tobacco plants (Nicotiana tabacum L. Havana 423) expressing the PRSV CP gene with the gus leader (GCP lines) or with the viral leader (5'CP lines) were obtained by Agrobacterium-mediated transformation. When the transgenic lines were analyzed by western blotting, the protein products of 36 kDa and 45 kDa reacting to PRSV CP antiserum were detected in the GCP lines and 5'CP lines, respectively. The presence of the CP gene in the transgenic tobacco was also confirmed by polymerase chain reaction (PCR) using primers specific to the CP gene. Analysis of segregation ratios in the R1 plants of four GCP lines and four 5'CP lines indicated that the CP gene in all of them was nuclearly inherited as a single dominant trait. R0 and R1 plants of the four GCP lines and four 5'CP lines were inoculated with tobacco etch virus (TEV), potato virus Y (PVY), or pepper mottle virus (PepMoV). The transgenic lines showed significant delay in symptom development and the severity of symptoms was attenuated. The GCP lines expressing the PRSV CP gene by the gus leader accumulated higher levels of CP and showed higher degrees of resistance than the 5'CP lines with the PRSV 5' leader. Our results indicate that the homologous viral leader does not enhance CP expression either in vitro or in vivo, nor does it provide better resistance in transgenic tobacco.
Keywords: Coat protein gene; Papaya ringspot virus; Potyvirus; Translation leader.
Introduction
Transgenic plants expressing the coat protein (CP) of a plant virus have been proved resistant to infection by the same or related viruses. The first illustration of CP-mediated resistance was reported by Powell-Abel et al. (1986). In that study the CP gene of tobacco mosaic virus (TMV) was transferred to tobacco, and symptom development was delayed in the transgenic plants when challenged with TMV. Since then, there have been numerous reports demonstrating that transgenic plants with viral CP genes are resistant to virus infection (Beachy et
al., 1990; Fitchen and Beachy, 1993). The level of resistance ranged from delay of symptom development to immunity, and the spectrum of resistance to viruses ranged from narrow to broad. The CP-mediated protection has provided an innovative method for control of plant viruses.
Papaya ringspot virus (PRSV) is a member of the genus Potyvirus, the largest group of plant viruses (Reichmann et al., 1992). Gene expression and genetic organization of the virus have been characterized previously (Yeh and Gonsalves, 1985; Yeh et al., 1992). The genome of PRSV contains one large ORF, which encodes a polyprotein of 383 kDa that is proteolytically processed into functional proteins. The destructive papaya ringspot disease caused by the virus is the major factor limiting the wide-scale planting of this fruit tree throughout papaya
3Corresponding author. Tel: 04-2877021; Fax: 04-2877585; E-mail: sdyeh@nchu.edu.tw