Bot. Bull. Acad. Sin. (2000) 41: 197-202
Lee and Chou Microcystins from cultured Microcystis aeruginosa
Isolation and identification of seven microcystins from a
cultured M.TN-2 strain of Microcystis aeruginosa
Tzong-Huei Lee1 and Hong-Nong Chou2,3
1Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan 11529
2Institute of Fisheries Science, National Taiwan University, Taipei, Taiwan 10617
(Received October 30, 1999; Accepted March 6, 2000)
Abstract. Seven microcystins (MCYSTs), three major and four minor ones, were isolated from a cultured Microcystis aeruginosa strain M.TN-2 isolated from an eel pond in southern Taiwan in the summer of 1993. Microcystins were separated by a column separation of LH-20 gel filtration chromatography, a subsequent silica gel flash column chromatography, and then purified by a reversed phase high performance liquid chromatography. MCYST-LR, -RR, -RA, -FR, -WR, [D-Asp3] MCYST-FR, and [D-Asp3] MCYST-WR were characterized and determined by amino acid analysis, MALDI-TOF mass spectrometry, 1H and DQF-COSY nuclear magnetic resonance spectroscopy and UV spectroscopy. Among them, [D-Asp3] MCYST-FR and [D-Asp3] MCYST-WR, two new compounds, were the desmethyl variants of MCYST-FR and MCYST-WR, respectively.
Keywords: Cyanobacteria; Hepatotoxin; Microcystin; Microcystis aeruginosa.
Introduction
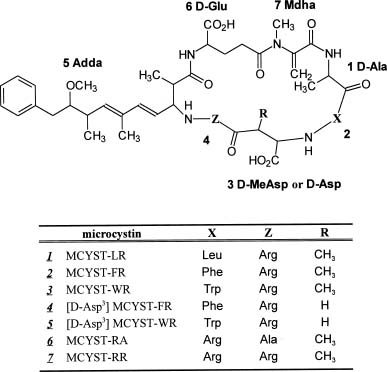
Toxic cyanobacteria have caused fatal poisoning of mammals, birds, and fish (Codd and Poon, 1988; Carmichael, 1992; Kotak et al., 1996; Tencalla and Dietrich, 1997), and may also have exerted adverse effects on human health (Gorham and Carmichael, 1988; Carmichael, 1994, 1997). Microcystins, a vast group of cyanobacterial toxins, are known to be hepatotoxic cyclic heptapeptides produced by some species within the genera of Microcystis (Shirai et al., 1991; Namikoshi et al., 1992; Luukkainen et al., 1994; Lee et al., 1998), Oscillotoria (Meriluoto et al., 1989; Luukkainen et al., 1993), Nostoc (Namikoshi et al., 1990; Sivonen et al., 1990; Beattie et al., 1998), and Anabaena (Harada et al., 1991a; Sivonen et al., 1992; Namikoshi et al., 1998). Toxic Microcystis species are widely distributed, mainly in the fresh water world, and form blooms frequently. The general structure of microcystins is composed of (2S, 3S, 8S, 9S)-3-amino-9-methoxy-2,6,8-trimethyl-10-phenyldeca-4,6-dienoic acid (Adda), g-linked D-glutamic acid (D-Glu), N-methyldehydroalanine (Mdha), and D-alanine (D-Ala). Four special amino acids are linked in sequential order followed by two variable L-amino acids with D-erythro-b-methylaspartic acid (D-MeAsp) inserted between them, forming a cyclic structure (Figure 1). There have been more than 60 different components isolated and determined. These compounds differ primarily in their L-amino acids at the number 2 and number 4 position of the
peptide ring skeleton, and in the presence or absence of the methyl group on D-MeAsp and/or Mdha of the 3rd and 7th amino acid, respectively (Namikoshi et al., 1992). L-amino acids in microcystins at the 2nd and 4th position are known to be arginine, leucine, tyrosine, phenylalanine, tryptophan and others. Among the identified microcystins, more than 70% of the structures contained L-Arg at the 4th position. Among these 7 cyclic-linked amino acids in microcystins, Adda has been thought of
3Corresponding author. Tel: +886-2-23639291-125; Fax: +886- 2-23629919; E-mail: unijohn@ccms.ntu.edu.tw
Figure 1. Structures of microcystins identified in this research.