Bot. Bull. Acad. Sin. (2000) 41: 219-223
Chang et al. Phalaenopsis and Doritis cpDNA inheritance patterns
RFLP and inheritance patterns of chloroplast DNA in
intergeneric hybrids of Phalaenopsis and Doritis
Song-Bin Chang1,2, Wen-Huei Chen1,2,3,4, Hong-Hwa Chen2,3, Yang-Ming Fu1, and Yih-Shyan Lin1
1Department of Horticulture, Taiwan Sugar Research Institute, Tainan, Taiwan 702, Republic of China
2Department of Biology, and 3Institute of Biotechnology, National Cheng Kung University, Tainan, Taiwan 701, Republic of China
(Received June 22, 1999; Accepted November 30, 1999)
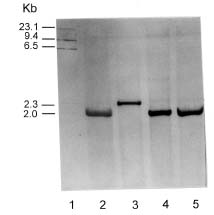
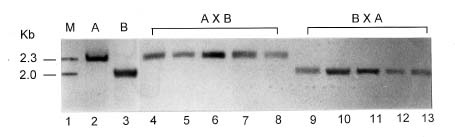
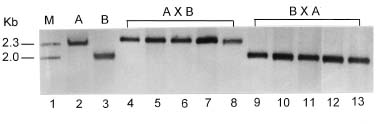
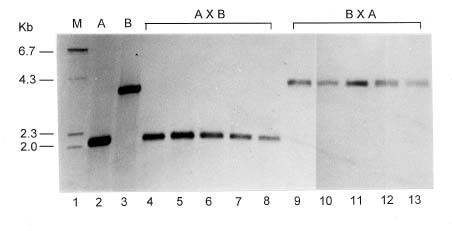
Abstract. The mode of inheritance of chloroplasts was analyzed using restriction fragment length polymorphism (RFLP) in both interspecific hybrids of Phalaenopsis and intergeneric hybrids of Phalaenopsis and Doritis. Chloroplast DNA digested with Dra I followed by hybridization with an rbcL probe revealed that Phalaenopsis amabilis, Phalaenopsis aphrodite, and Phalaenopsis stuartiana, which belong to the taxonomic section PHALAENOPSIS, have the same size 2.0-kb fragment. Both Phalaenopsis mannii and Phalaenopsis amboinensis have a 2.3-kb fragment, while Doritis pulcherrima has a 3.5-kb fragment. In both interspecific and intergeneric hybrids, maternal inheritance of the chloroplast genome was detected. The hybrids of both reciprocal crosses (A x B and B x A) are registered with the same hybrid names in Sander's List of Orchid Hybrids at the Royal Horticultural Society, despite harboring chloroplast DNA from different parents. These results suggest that the chloroplast DNA can be used as a marker for identification of parentship and for phylogenetic studies of taxonomy.
Keywords: cpDNA inheritance pattern; Doritis; Phalaenopsis; RFLP.
Introduction
Phalaenopsis spp., and Doritis spp. belong to Tribes Vandeae and Orchidaceae, respectively. About 45 wild species of Phalaenopsis are distributed in Asia in a region 23° north and south of the equator (Sweet, 1980). A large collection of various Phalaenopsis spp. is maintained at the Taiwan Sugar Research Institute (TSRI) including 34 wild species and 1,239 superior hybrids (Chen and Wang, 1996). The botanic and horticultural characteristics of these plants have been analyzed for breeding purposes.
Unlike nuclear genes, the inheritance pattern of organelle genes varies greatly among different organisms (Birky, 1995). So far, the inheritance of cytoplasmic organelles in Orchidaceae has been studied only cytogenetically using DNA fluorochrome 4', 6'-diamidino-2-phenyl indole (DAPI) in conjunction with epifluorescence microscopy (Corriveau and Coleman, 1988). However, no molecular analyses have been carried out in Phalaenopsis. Here we present that the maternal inheritance pattern of chloroplast DNA (cpDNA) was detected in hybrids of Phalaenopsis and Doritis by using RFLPs visualized with a rbcL probe.
Materials and Methods
Plant Materials
Intraspecific and interspecific crosses of Phalaenopsis and intergeneric crosses between Phalaenopsis and Doritis were performed and maintained in the greenhouse of TSRI (Table 1). Six wild species of Phalaenopsis and one wild species of Doritis were used in these experiments, including P. amabilis (L.) Blume, P. amboinensis J. J. Smith, P. aphrodite Rchb. F., P. equestris (Schauer) Rchb. f., P. mannii Rchb. f., P. stuartiana Rchb. f., and D. pulcherrima Lindl. Among them, P. amabilis (L.) Blume, P. aphrodite Rchb. F., and P. stuartiana Rchb. F. belong to the same taxonomic section PHALAENOPSIS (Fu et al., 1997). One intraspecific hybrid between P.
Table 1. Plant materials used in crosses.
Intraspecific crosses
P. equestris `W9-55' x P. equestris `W9-57'
P. equestris `W9-57' x P. equestris `W9-55'
Interspecific crosses
P. amabilis `W1-2' x P. amboinensis `W2-2'
P. amboinensis `W2-2' x P. amabilis `W1-2'
P. mannii `W25-1' x P. stuartiana `W40-5'
P. stuartiana `W40-5' x P. mannii `W25-1'
Intergeneric crosses
P. equestris `W9' x D. pulcherrima `W46-26'
D. pulcherrima `W46-26' x P. equestris `W9'
4Corresponding author. Tel: 886-6-289-1853; Fax: 886-6-268-5425; E-mail: a08539@taisugar.com.tw