Bot. Bull. Acad. Sin. (2000) 41: 231-236
Chung Spatial genetic structure in Hemerocallis
Spatial genetic structure in three populations of Hemerocallis hakuunensis (Liliaceae)
Myong Gi Chung
Department of Biology, Gyeongsang National University, Chinju 660-701, The Republic of Korea
(Received May 21, 1999; Accepted December 24, 1999)
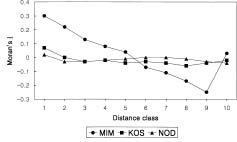
Abstract. Ninety-nine, 75, and 71 individuals were mapped and leaf samples were collected from three populations (10 × 15-m plot [MIM], 40 × 50-m plot [KOS], and 15 × 20-m plot [NOD]) of Hemerocallis hakuunensis to determine if spatial genetic structure existed in the three populations in terms of their ecological characteristics. A substantial spatial genetic structuring was found in MIM, whereas a weak structure was observed in NOD and KOS. Moran's I values were significantly different from the expected value (-0.010) in 137 (52.7%) of 260 cases in MIM, whereas significant Moran's I values were observed in 25 (10.4%) of 240 cases and 55 (20.4%) of 270 cases in NOD and KOS, respectively. The approximate minimum patch width also varied among the three populations: NOD = 3 m, MIM = 5 m, and KOS = 10 m. The differences in patch width may result from the differences in density, colonization history, thinning processes or any of a number of other factors among populations.
Keywords: Allozymes; Density; Hemerocallis hakuunensis; Moran's I; Spatial autocorrelation.
Introduction
During the past decade, spatial genetic structure has been quantified using spatial autocorrelation statistics using allozymes as genetic markers to understand the evolutionary dynamics of plant populations (e.g., Berg and Hamrick, 1995; Chung and Epperson, 1999; Dewey and Heywood, 1988; Epperson and Clegg, 1986; Perry and Knowles, 1991; Schnabel and Hamrick, 1990). These studies showed that individuals are not likely to be randomly distributed owing to factors such as limited seed and pollen dispersal, isolation in small patches, differential mortality, and microenvironmental selection. In addition to these factors, differences in density, pollinator behaviors, topography, and colonization history could result in a different genetic architecture in plant populations of a species (e.g., Chung et al., 1998; Knowles et al., 1992; Young and Merriam, 1994). All these aspects should be considered in studies of natural plant populations to gain insights into their maintenance mechanisms of genetic diversity.
Hemerocallis hakuunensis Nakai is commonly found in grasslands of mountainous areas, under pineoak forests on the hillsides, and under disturbed orchards of chestnut trees of the southern, central, and northwestern Korean Peninsula (Chung and Kang, 1994a). Individuals of the species have 5 to 16 flowers on the branched inflorescence. Flowers are orange-yellow and are usually visited by bees, bumblebees, and flies. They start to open before sunrise and remain open until the afternoon (Chung, pers. obs.). The species has no specialized mechanisms for seed dispersal (Chung, pers. obs.).
In this study, spatial autocorrelation using allozyme markers was conducted in three populations of H. hakuunensis to determine whether differences in spatial genetic structure existed in populations under different ecological conditions.
Materials and Methods
In August 1995 and August 1997, 99, 75, and 71 individuals were mapped and leaf samples were collected within a 10 × 15-m plot (Mich'eon-myeon, Chinju-shi, Prov. Gyeongsangnam-do, hereafter referred to as MIM), a 40 × 50-m plot (Sangri-myeon, Kosung-gun, Prov. Gyeongsangnam-do, hereafter referred to as KOS), and a 15 × 20-m plot (Nogodan, Mts. Chiri, Prov. Chollanam-do, hereafter referred to as NOD), respectively. Ecological characteristics of the three populations are summarized in Table 1. Leaf samples were placed in plastic bags wrapped with a wet paper towel and stored on ice to prevent protein denaturation prior to returning to the laboratory, where they were stored at 4°C until protein extraction.
Leaf samples were cut finely, and crushed with a mortar and pestle. A phosphatepolyvinylpyrrolidone extraction buffer (Mitton et al., 1979) was added to the leaf samples to facilitate crushing and to aid enzyme stabilization. The cellular extract was absorbed onto 4 × 6-mm Whatman 3MM chromatography paper wicks, which were then stored at -70°C until needed. Electrophoresis was performed using 11% starch gels. Fourteen putative loci were resolved from eight enzyme systems using three gel/electrode buffer combinations. Two Poulik buffer systems were used: a modification (Haufler, 1985) of Soltis et al. (1983) "system 6" was used to resolve leucine aminopeptidase (LAP), fluorescent esterase (FE) and a modi
Fax: +82-591-754-0086; E-mail: mgchung@nongae.gsnu.ac.kr