Bot. Bull. Acad. Sin. (2000) 41: 305-314
Abd El-Ghani Vegetation composition of Egyptian inland saltmarshes
Vegetation composition of Egyptian inland saltmarshes
Monier M. Abd El-Ghani
The Herbarium, Faculty of Science, Cairo University, Giza 12613, Egypt
(Received July 6, 1999; Accepted May 11, 2000)
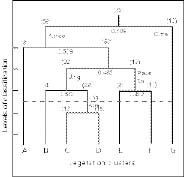
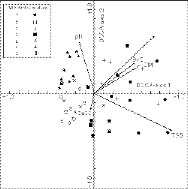
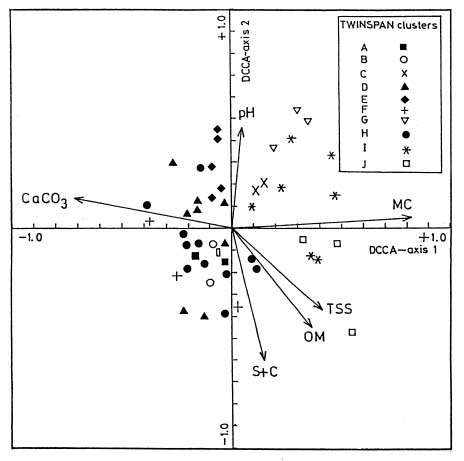
Abstract. The vegetation-environment relationships in the inland saltmarshes of two geographically distant oases of the Western Desert of Egypt, Siwa, and Dakhla Oases, are described. Two data sets (25 species × 68 stands for Siwa Oasis and 29 species × 89 stands for Dakhla Oasis) were analysed, using multivariate procedures, i.e., two-way indicator species analysis (TWINSPAN), detrended correspondence analysis (DCA), and detrended canonical correspondence analysis (DCCA), to produce a classification of plant communities in the studied areas, and to examine the relationships of these plant communities to certain edaphic factors, namely soil reaction, total soluble salts, calcium carbonate, organic matter, moisture content, and fine fractions. Twelve halophytic plant communities linked to two main habitats (wet-moist and dry-mesic) were identified. Alhagi graecorum, Tamarix nilotica, Cressa cretica, Juncus rigidus and Phragmites australis were the most common in the two oases. Whereas communities of Cyperus laevigatus, Suaeda aegyptiaca, Suaeda vermiculata, Typha domingensis, and Aeluropus lagopoides are recorded from the Dakhla Oasis, Cladium mariscus and Arthrocnemum macrostachyum communities are recorded from the Siwa Oasis. The most important edaphic variables affecting the distribution and structure of the plant communities are salinity, moisture content and fine fractions, nevertheless CaCO3 content seems to be more effective in the Dakhla Oasis. A new agricultural strategy that minimizes the increase of salinized lands is called for.
Keywords: Egypt; Halophytes; Multivariate analysis; Oases; Phytosociology; Saltmarsh vegetation; Secondary salines.
Introduction
Saline lands are widely distributed globally and make up about 10% of the Earth's terrestrial surface (O'Leary and Glenn, 1994). Compared to studies of coastal marshes, little attention has been paid to inland saline landscapes (Adam, 1990; Krüger and Peinemann, 1996). The inland saltmarshes of the Egypt's Western Desert are found in the form of Sabkhas (Zahran, 1982) around the lakes, springs and wells of the oases, e.g., Siwa, Dakhla, Kharga, Bahariya and Farafra, and depressions, e.g., Qattara, Wadi El-Natrun, and El-Faiyum (Figure 1). Being lower in altitude than the surrounding territories, the inland saltmarshes are characterized by a shallow underground water table. In certain instances, the underground water is exposed, forming lakes of brackish or saline water (Zahran and Girgis, 1970; Zahran, 1972). The formation of these salines is due to the uncontrolled spilling of water and flooding of the plains or to the water table, which is near to the ground (Migahid et al., 1960). Under the severe arid conditions of the oases and the lack of a drainage system, flooding of the soil with slightly saline artesian water rapidly increases its salinity. In contrast to the littoral saltmarshes, these salines can be considered as secondary. The vegetation has a patchy structure, with different patches containing different species (or sometimes one species) and even different growth forms (Abu-
Ziada, 1980; El-Hadidi, 1993). Despite the low number of halophytes in Egypt, with 80 terrestrial plant species from 17 families (Batanouny and Abo Sitta, 1977), they constitute the principal vegetation of extensive areas in the country. The halophytic flora is poor, being composed mainly of perennial grasses, rushes, (dwarf-) shrubs, and some annuals which are associated with saline environments, e.g., Frankenia pulverulenta L., Lotus corniculatus L., Solanum nigrum L., Asphodelus tenuifolius Cav., Bassia muricata (L.) Asch., Anagallis arvensis L. (s.l.), Conyza bonariensis (L.) Cronquist and Ambrosia maritima L.
Although the autecology, synecology and ecophysiology of several coastal saltmarshes of the Western Desert have been dealt with in a large number of publications (e.g. Ayyad and El-Ghareeb, 1974, 1982; Fahmy, 1986; Shaltout and El-Ghareeb, 1992; Zahran et al., 1996), studies identifying the major environmental factors associated with vegetation patterns in the inland saltmarshes are scarce. What studies there are have attempted to elucidate, by various means, factors causing the differences in communities both within and between marshes. However, objective methodology and quantitative procedures have been applied in the present study using techniques involving classification and ordination. Recently, some studies in different parts of Egypt (Abd El-Ghani, 1998, 1999; Dargie and El-Demerdash, 1991; Moustafa and Zaghloul, 1996; Shaltout et al., 1995; Springuel et al., 1997), based on a multivariate approach to plant community analysis, were carried out.
E-mail: elghani@yahoo.com