Bot. Bull. Acad. Sin. (2001) 42: 85-92
Chang et al. Rice class I LMM HSP gene, Oshsp16.9C
Isolation and characterization of the third gene
encoding a
16.9 kDa class I low-molecular-mass heat shock protein, Oshsp
16.9C, in rice
Pi-Fang Linda Chang1,3, Chung-Yi Huang2,3, Fa-Cheng Chang2, 4, Tong-Seung Tseng2, 5, Wan-Chi Lin2, 6, and Chu-Yung Lin2,*
1Department of Plant Pathology, National Chung Hsing University, Taichung, Taiwan 402, Republic of China
2Department of Botany, National Taiwan University, Taipei, Taiwan 106, Republic of China
(Received May 1, 2000; Accepted September 6, 2000)
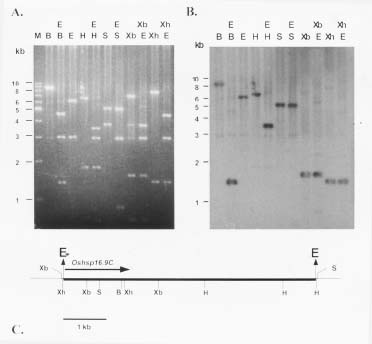
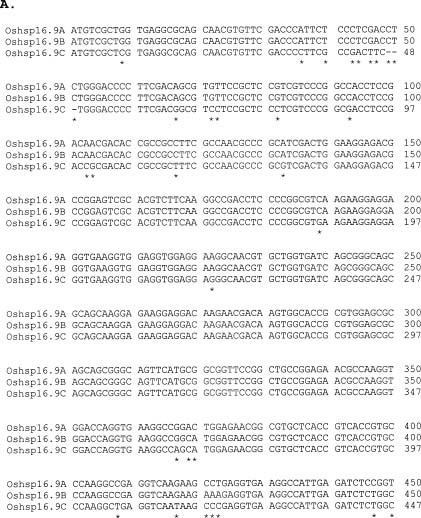
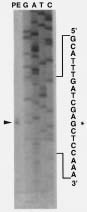
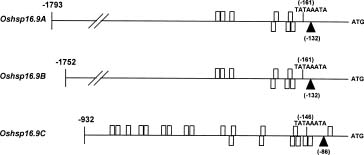
Abstract. Using rice cDNA pTS1 encoding a 16.9 kDa heat shock protein (HSP) as a probe for Southern hybridization analysis, we observed five prominent bands of 9.5, 5.9, 3.4, 2.5, and 1.7 kb in the Eco RI digests of rice genomic DNA and found them to contain six individual genes. The 5.9 kb DNA fragment was further digested with Hind III to generate three fragments of 3.5, 1.7 and 0.7 kb, and we found, using pTS1 cDNA as a probe, the 3.5 kb fragment contained a putative low-molecular-mass (LMM) HSP gene. The DNA sequencing of 3.5 kb fragment revealed encoding of a presumptive 16.9 kDa HSP (149 amino acid residues) with predicted pI value of 6.42. The nucleotide sequence of this gene was highly homologous to the coding regions of two rice class I LMM HSP genes, Oshsp 16.9A and Oshsp16.9B, sharing 93.1% and 94.3% sequence identity, respectively, as published previously in our laboratory. The deduced amino acid sequence of this gene is similar to those of the Oshsp16.9A and Oshsp16.9B genes with a difference of only 11 and 10 amino acids, respectively. It was hence designated as Oshsp16.9C (accession no. U81385). We used 3' UTRs (untranslated regions) for analysis of Oshsp16.9C gene expression since the 3' UTRs of these three genes showed very low sequence homology.
Keywords: Heat shock protein; Heat shock gene; Low-molecular-mass heat shock protein; Oryza sativa L.
Abbreviations: HS, heat shock; HSP, heat shock protein; LMM, low-molecular-mass; UTR, untranslated region.
Introduction
Heat shock proteins (HSPs) have been induced during thermal stress in all organisms ever examined, ranging from bacteria to human beings (Schlesinger et al., 1982), and they appear to be involved in thermoprotection (Lin et al., 1984; Chou et al., 1989; Krishnan et al., 1989; Vierling, 1991; Jinn et al., 1997). The HSPs are usually divided into high-molecular-mass (HMM) proteins of more than 30 kDa and low-molecular-mass (LMM) proteins of about 17 to 28 kDa (Lindquist and Craig, 1988; Vierling, 1991). In contrast to animal systems, plants synthesize more abundant LMM HSPs than HMM HSPs. The LMM HSPs superfamily is unusually complex, consisting of at least five gene families (LaFayette et al., 1996; Waters et al., 1996). The role
of LMM HSP in heat stress is not completely clear yet. However, the sequence conservation of genomic and cDNA clones of plant LMM HSP genes isolated and characterized from a number of species suggests that they may play an important role in plants coping with HS.
We have been studying the physiological function of class I LMM HSPs in soybean and rice (Lin et al., 1984; Chou et al., 1989; Jinn et al., 1989; Jinn et al., 1993; Jinn et al., 1995; Yeh et al., 1995; Jinn et al., 1997; Yeh et al., 1997). We have isolated and characterized three cDNA clones: pTS1 (encoding a 16.9 kDa HSP, Tseng et al., 1992), pTS3 (encoding a 17.3 kDa HSP, Tseng et al., 1992), and pYL (encoding a 18.0 kDa HSP, Lee et al., 1995), and also five genomic clones, Oshsp16.9A, Oshsp16.9B, Oshsp18.0, Oshsp17.3, and Oshsp17.7 of rice class I LMM HSPs (Tzeng et al., 1992; 1993; Guan et al., 1998, respectively). The Oshsp16.9A contains the sequence of pTS1 cDNA. The three cDNA clones are highly homologous in their sequences except for the 3' untranslated regions (UTRs), which show a low degree of homology. Because of the abundance and complexity of these proteins, we have tried to isolate and characterize additional genes for rice class I LMM HSPs for the purpose of studying their differences in gene expression under heat stress. So far, we have isolated and characterized all the genes for rice class I LMM
3Equally contributed.
4Present address: Center of Genetic Engineering, National Chung Hsing University, Taichung, Taiwan 402, Republic of China.
5Present address: Department of Plant Biology, University of Minnesota, St. Paul, MN 55108-1095, USA.
6Present address: Institute of Bioagricultural Sciences, Academia Sinica, Taipei, Taiwan 115, Republic of China.
*Corresponding author. Tel: 886-2-2363-0231 ext. 2675; Fax: 886-2-2363-8598; E-mail: chuyung@ccms.ntu.edu.tw