Bot. Bull. Acad. Sin. (2001) 42: 109-114
Wei et al. Maize lines analysed by GISH
Comparative analyses of disease resistant and nonresistant lines from maize × Zea diploperennis by GISH
Wen-Hui Wei1, Rui Qin1, Yun-Chun Song1,*, Le-Qun Guo2, and Ming-Guang Gu2
1The Key Laboratory of MOE for Plant Developmental Biology, College of Life Sciences, Wuhan University, Wuhan Hubei 430072, The People's Republic of China
2Institute of Genetics, Chinese Academy of Sciences, Beijing 100101, The People's Republic of China
(Received April 27, 2000; Accepted October 9, 2000)
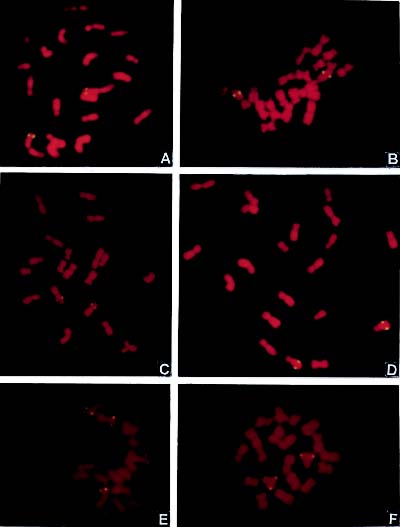
Abstract. After a cross between maize inbred line Lu9 and Zea diploperennis (DP) and a backcross between their F1 and Lu9, the BC1 was reproduced with parthenogenesis induced by chemicals. In this study a2-(1) and a2-(6), the two tested, stable alloplasmic sister lines, were obtained through selecting and selfing of the parthenogenetical progenies for over ten generations. The line a2-(1) showed DP characters, such as resistance to Helminthosporium turcium Pass, H. maydis Nisik, and H. carbonum ULLstrup while a2-(6) exhibited no such resistance. The introgressed DP DNA segments were successfully detected, and their physical location on chromosomes were compared in these two lines by genomic in situ hybridization (GISH). In line a2-(1), the hybridization signals were located on the long arms of chromosomes 1, 2 and 8, and on the short arm of chromosome 6; in line a2-(6), they were located only on the long arms of chromosomes 1, 2 and 8, with no signal on chromosome 6 at all. The distribution of introgressed DP segments on chromosomes, reasons for differences of disease resistance between these two lines, and the existence of resistance genes in the introgressed segments are discussed.
Keywords: Alien introgressed segments; Disease resistance genes; Genomic in situ hybridization (GISH); Maize; Zea diploperennis.
Introduction
Many fine crop varieties showing high yield, quality, and disease resistance have been obtained by distant hybridization, including maize (Guo et al., 1997; 1998), rice (Mohan et al., 1994), barley (Pickering et al., 1997), wheat (Schwarzacher et al., 1992), onion (Keller et al., 1996), and their wild relatives. Zea diploperennis (DP) found by Iltis et al. (1979) is the immune source for several maize viruses, and it is also resistant to maize Helminthosporium turcium Pass, H. maydis Nisik, H. carbonum ULLstrup, foliar and root pathogens as well as insect pests such as corn earworms, stalk borers, and rootworms. In addition, DP has stress tolerance to drought, excess water, and low temperature (Nault et al., 1981). Guo et al. (1997; 1998) crossed maize inbred lines Lu9 and 330 with Zea diploperennis (DP) and obtained several maize lines showing DP characters, such as disease resistance, stress resistance, high yield, and quality. Whether the DP DNA segments were steadily introgressed into the genome of these lines or notand if so, where such segments are locatedis still unknown.
Plant breeders are interested in detecting the alien DNA segments in distant hybrids. The technique of genomic in situ hybridization (GISH) has been widely used in the
analyses of genomic construction and the detection of alien chromatin (Durnam et al., 1985; Pinkel et al., 1986; Schwarzacher et al., 1989; Barre et al., 1998; Humphreys et al., 1998; Khrustaleva and Kik, 1998; Kamstra et al., 1999; Liu et al., 2000; Zhang et al., 2000). In this paper, we describe the detection and location of the introgressed alien segments in two alloplasmic pure lines from inbred Lu9 × DP, one disease resistant and the other susceptible.
Materials and Methods
Plant Materials
The materials were obtained by Guo et al. (1997; 1998). Maize inbred line Lu9 was crossed with Zea diploperennis (DP) in 1985. Subsequently the F1 was backcrossed with its maize parent and produced BC1 in 1986. BC1 was reproduced parthenogenetically by treatment with chemicals in 1987. Guo et al. further selected and bred maize lines of high yield and quality with some DP characters, such as disease and stress resistance, by selfing for over ten generations, eventually obtaining several stable alloplasmic lines. Two stable, pure lines a2-(1) and a2-(6) and their parents, maize inbred lines Lu9 and DP, were chosen as the tested materials. According to inoculated tests in field and greenhouse (Guo et al., 1997), line a2-(1) was resistant to maize Helminthosporium turcium Pass, H.
*Corresponding author. E-mail: ycsong@whu.edu.cn