Bot. Bull. Acad. Sin. (2001) 42: 131-139
Hwang and Chen Nutrients and growth in Kandelia candel
Effects of ammonium, phosphate, and salinity on growth, gas exchange characteristics, and ionic contents of seedlings of mangrove Kandelia candel (L.) Druce.
Yuan-Hsun Hwang* and Shuh-Chun Chen
Institute of Botany, Academia Sinica, Taipei 115, Taiwan, Republic of China
(Received April 17, 2000; Accepted August 24, 2000)
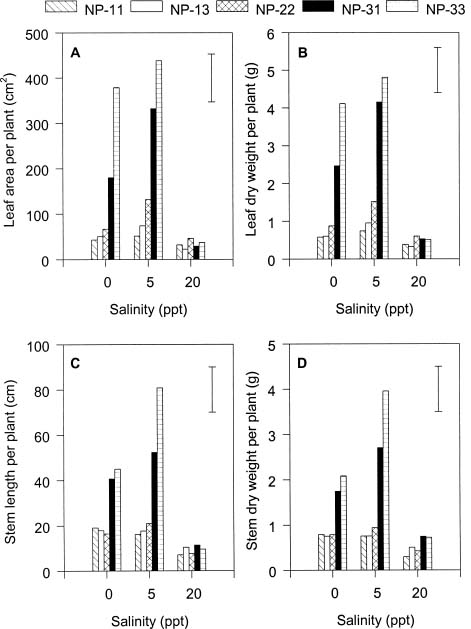
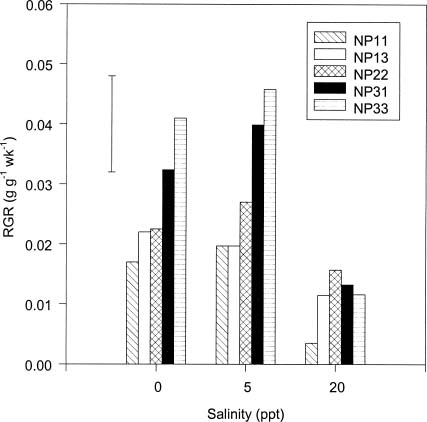
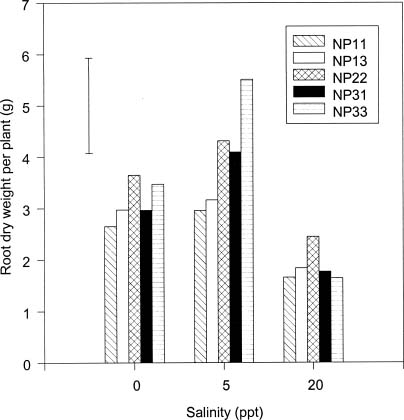
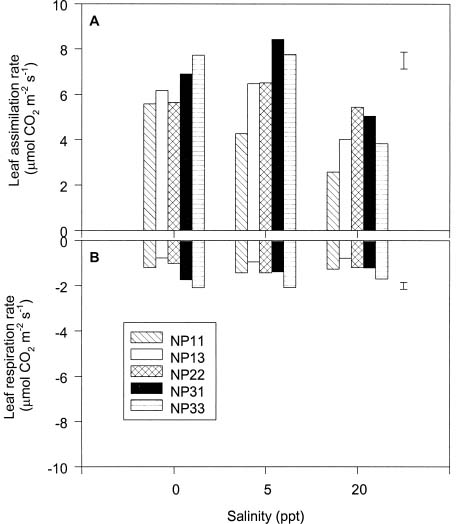
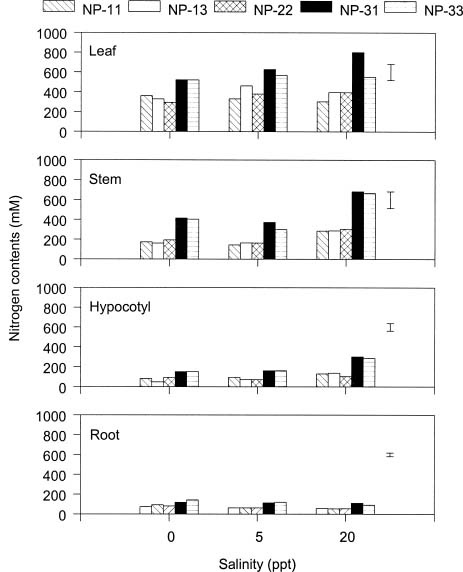
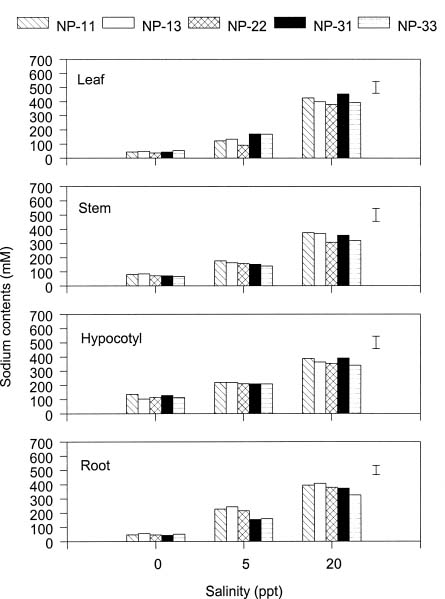
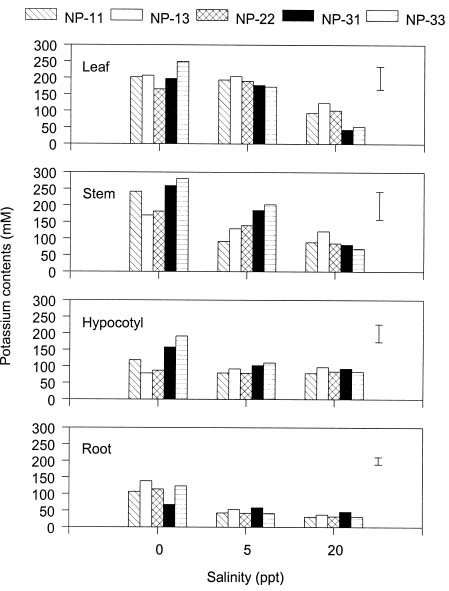
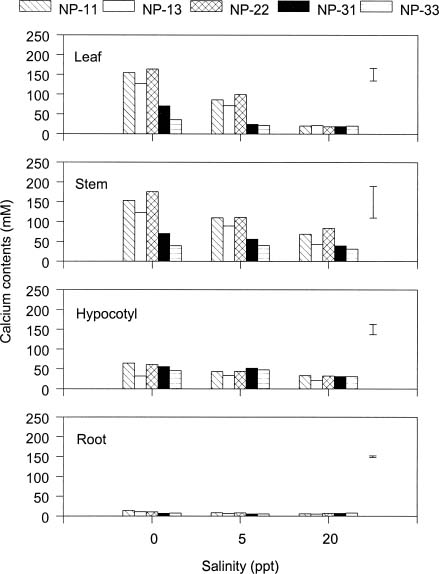
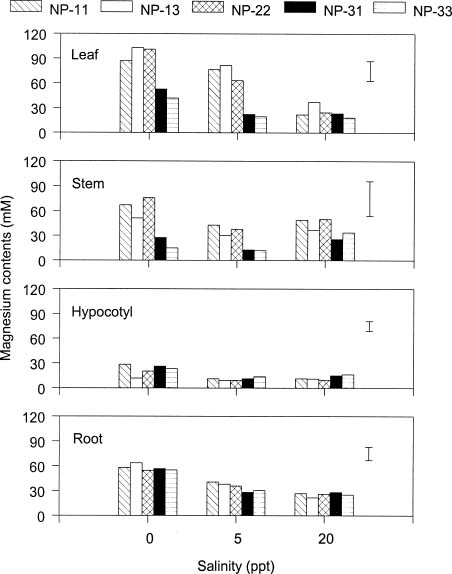
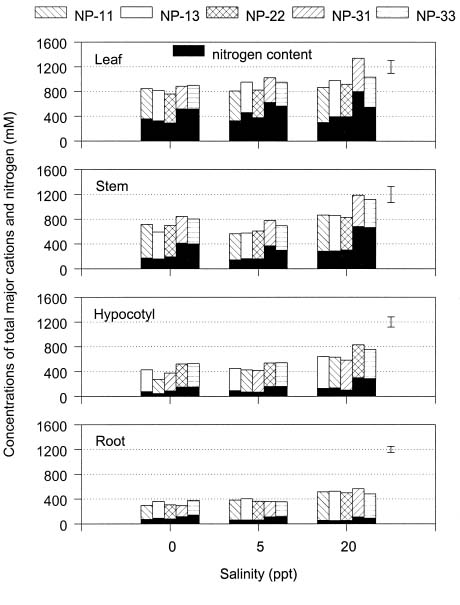
Abstract. The effects of ammonium, phosphate, and salinity on growth, gas exchange characteristics, and ionic content of seedlings of mangrove Kandelia candel (L.) Druce were investigated in solution culture experiments over seven months. High salinity at 20 ppt NaCl greatly reduced dry matter accumulation in shoots and roots and leaf CO2 assimilation rate. Addition of ammonium-nitrogen (2 mM) significantly increased the growth of shoots and roots, leaf CO2 assimilation rate, and respiration rate at low salinity treatments. Phosphate amendment alone did not affect the growth of plants. However, when ammonium-nitrogen was also added, phosphate improved the growth of plants, leaf CO2 assimilation rate, and respiration rate in low salinity treatments. Kandelia candel tended to maintain a constant cation concentration in tissues of leaves and roots at low salinity conditions. Potassium was the major cation in the tissues at the low salinity; however, it was replaced by sodium as the salinity of the culture solution was increased. Kandelia candel absorbed ammonium-nitrogen luxuriously at all salinity treatments and some of the ammonium accumulated in the tissues could be part of cations as the osmotic inorganic solute. Reasons for the difference in optimal growth salinity for K. candel between porewater in the field and the culture solution in the greenhouse are discussed.
Keywords: Ammonium; Growth; Kandelia candel; Mangrove; Nitrogen; Nutrients; Phosphate; Salinity.
Introduction
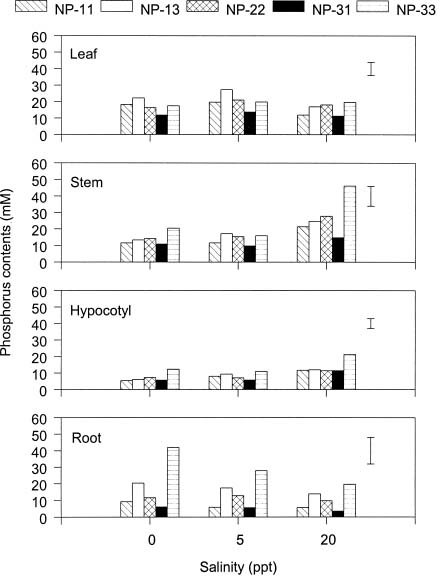
The interstitial water and sediment of the stunted mangrove forest in the Chuwei mangrove wetland in Taipei, Taiwan are deficient in both nitrogen and phosphorus (Hwang, 1983; Hwang and Hsu, 1996; Kao and Chang, 1998). A recent field fertilization study indicated that nitrogen is the major nutrient limiting the growth of the stunted mangrove Kandelia candel (L.) Druce (Rhizophoraceae) in this wetland (Chen, 1998). The fact that stunted mangrove plants growing in low phosphorus environments did not respond to the addition of phosphorus fertilizer seems unusual, considering results from other mangrove forests in northern Australia (Boto and Wellington, 1983) and in Belize (Feller, 1995). Both of those studies indicated that phosphorus was the major nutrient limiting the growth of Rhizophora spp. in the interior, higher elevation areas of the forest.
The availability of essential nutrients, especially phosphorus ion, in waterlogged mangrove soils is largely controlled by the redox potential of the sediment (Patrick and Mahapatra, 1968; Ponnamperuma, 1972). Thus, the lack of apparent response to phosphorus fertilization in K. candel (Chen, 1998) may indicate that K. candel is not phosphorus limited or that the added phosphorus is un
available for uptake due to precipitation under oxidizing conditions. Additional investigation is required to evaluate these possibilities.
Alternatively, the comparative importance of nitrogen-limitation on the growth of K. candel may be related to salinity. Salinity has long been recognized as an important factor regulating growth and distribution of mangroves (Ball, 1988; Lin and Sternberg, 1993; Ball and Pidsley, 1995). Previous greenhouse studies on the growth of K. candel seedlings indicated that the optimal salinity for growth was 85 mM NaCl (5 ppt) and that growth was inhibited at salinities above 340 mM NaCl (20 ppt) (Hwang and Chen, 1995). In contrast, the porewater salinity in the Chuwei mangrove swamp was between 25 and 30 ppt (Hwang, 1983; Hwang and Hsu, 1996; Chen, 1998). These observations suggest that the in situ salinity in the Chuwei mangrove swamp may limit the growth of K. candel. The accumulation of nitrogenous metabolites in angiosperm halophytes is well documented and may affect the ability of such plants to grow in high salinity conditions (Storey and Wyn Jones, 1975; Popp et al., 1984). Thus the nitrogen fertilization-induced growth increase observed in K. candel under high salinity conditions may reflect improved salt tolerance.
The purpose of this study was to examine the growth responses and tissue nutrient contents of Kandelia candel to variations in salinity, nitrogen and phosphate under greenhouse culture conditions.
*Corresponding author. Fax: 886-2-27827954; E-mail: bohwang@ccvax.sinica.edu.tw