Bot. Bull. Acad. Sin. (2001) 42: 167-171
Jing et al. Mapping fertility-restoring genes of rice WA cytoplasmic male sterility using SSLP markers
Mapping fertility-restoring genes of rice WA cytoplasmic male sterility using SSLP markers
Runchun Jing, Xiaoming Li, Ping Yi, and Yingguo Zhu*
The Key Laboratory of MOE for Plant Developmental Biology and Institute of Genetics, College of Life Sciences, Wuhan University, Wuhan, 430072, P.R. China
(Received August 17, 2000; Accepted November 16, 2000)
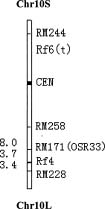
Abstract. Rice wild abortive (WA) type cytoplasmic male sterility (CMS) is commercially used for production of hybrid seeds in China. Two fertility restorer genes in the CMS-WA system were detected with SSLP markers in this study. An F2 population consisting of 210 excessive sterile individuals from a cross between Zhenshan 97A and a strong restorer line IR24 was used for mapping of Rf4. The genetic distance from Rf4 locus to RM171 (OSR33) and RM228 on long arm of chromosome 10 was 3.7 cM and 3.4 cM, respectively, which were the two closest SSLP markers flanking the Rf4 locus. The two SSLP markers gave promise of application in molecular marker-assisted selection (MAS) for fertility restorer lines of the CMS-WA system. RM244, another SSLP marker on the short arm of chromosome 10, was found to be linked with a fertility restorer locus in an F2 population consisting of 30 excessive sterile individuals from a cross between Zhenshan 97A and a weak restorer line IR64. The genetic relationship among fertility restorer genes Rf1, Rf4 and Rf5(t) for three rice CMS system is also discussed.
Keywords: Fertility restorer gene; Genetic linkage map; Molecular marker-assisted selection; Oryza sativa L.; SSLP markers.
Introduction
Plant cytoplasmic male sterility (CMS) caused by lesion or rearrangement of mitochondrial genome is unable to produce functional pollens. But CMS can be restored by nuclear genes. Therefore, the CMS systems are widely used for hybrid seed production. In rice, hybrid rice varieties developed based on wild abortive (WA) type CMS accounted for approximately 90% of hybrid rice in China (Yuan, 1992) and inheritance of fertility restoration for WA type CMS has been extensively studied. Most researchers conclude that the fertility restoration of WA type CMS is controlled by two independent nuclear loci. Besides CMS-WA, other CMS systems have also been developed and their fertility restoring genes were termed. Rf1, Rf2, Rf3 and Rf4 were the terms for CMS-BT, CMS-L and CMS-WA, respectively (Shinjyo, 1975; Zhang et al., 1997; Shinjyo and Sato, 1994). These genes were mapped on different chromosomes based on trisomic and RFLP analyses. Rf1 and Rf2 genes were mapped on chromosomes 10 and 2 by trisomic analyses, respectively (Shinjyo, 1975; Shinjyo and Sato, 1994). Rf3 and Rf4 were located on chromosomes 1 and 10 by trisomic and RAPD/RFLP analyses (Bharaj et al., 1995; Zhang et al., 1997; Yao
et al., 1997). Rf3 was flanked by RFLP markers RG458/RG140 and RG532, which had a genetic distance of 3.8 cM (Zhang et al., 1997). Rf4 was located in a 22.4 interval between G4003 and C234 (Yao et al., 1997). Tan et al. (1998) reported that one QTL, tightly linked to RFLP marker C1361 on chromosome 10, explained 71.5% of phenotypic variance (Tan et al., 1998). Huang et al. (2000) mapped a nuclear fertility restorer gene Rf5(t) on chromosome 10 for Honglian (HL) type CMS, another CMS type applied in commercial hybrid rice seed production in China (Huang et al., 2000).
Simple Sequence Length Polymorphism (SSLP) provides a new tool for gene mapping and marker-assisted selection (MAS). SSLPs are the results of variable numbers of repeated units within the microsatellite structure and are widely distributed in the rice genome (McCouch et al., 1997). SSLP can be easily and economically analyzed by Polymerase Chain Reaction (PCR) and generally behaves as a co-dominant marker, which often detects higher levels of allelic variation than RFLP or RAPD markers. SSLP markers are being successfully applied in fingerprinting and variety identification, gene and QTL mapping, and marker-assisted selection (MAS) in plants.
The studies reported in this paper were to determine locations of Rf loci in the rice genome based on linkage maps of SSLP markers (Akagi et al., 1996a; Panaud et al., 1996; Chen et al., 1997; Temnykh et al., 2000).
*Corresponding author. Tel: +86-27-87684560; Fax: +86-27-87882661; E-mail: ygzhu@hotmail.com