Bot. Bull. Acad. Sin. (2001) 42: 193-199
Liu et al. Adsorption protein of filamentous phage
Adsorption protein of filamentous bacteriophage fXo from Xanthomonas oryzae
Tzu-Jun Liu1, Bih-Yuh You2, Tsai-Tien Tseng3, Nien-Tsung Lin4, Ming-Te Yang, and Yi-Hsiung Tseng*
Institute of Molecular Biology, National Chung Hsing University, Taichung 402, Taiwan
(Received June 28, 2000; Accepted January 19, 2001)
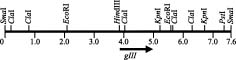
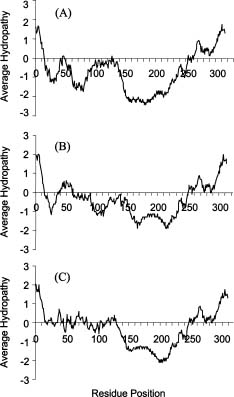
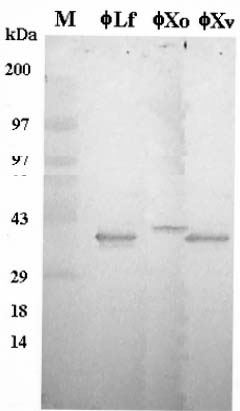
Abstract. fXo, Xf, fLf, fXv, and Cf are filamentous bacteriophages isolated in Taiwan. Both fXo and Xf specifically infect Xanthomonas oryzae pv. oryzae, and fLf, fXv, and Cf specifically infect X. campestris pv. campestris, X. campestris pv. vesicatoria, and X. campestris pv. citri, respectively. In this study, the fXo gene III (gIII) encoding the adsorption protein (pIII) was cloned by probing with the fLf gIII. Sequence analysis revealed that the fXo gIII is 1,023-nt long and able to encode a pre-protein of 340 aa (35,337 Da), with structural features typical of filamentous phage adsorption proteins: an N-terminal signal sequence (18 aa), a central region (90 aa) containing 38 glycine, 29 aspartic acid and 19 histidine residues, and a C-terminal membrane-anchoring domain (17 aa). The fXo pIII purified from the phage particles migrated as a 42-kDa band in SDS-polyacrylamide gel, which is substantially larger than that deduced from the nucleotide sequence, presumably due to the presence of the long stretch of charged residues in the central region. The fXo pIII could cross-react with the antiserum specific to fLf pIII, which also cross-reacts with fXv pIII. Like the situations in fLf and fXv, the pIIIs of fLf and fXo are also interchangeable. The gIII and the flanking regions of fLf, fXv and fXo are highly homologous and similar in size, but fXo has a genome (7.6 kb) larger than that of fLf (6.0 kb) and fXv (6.4 kb), suggesting that fXo is able to accommodate more genes and/or have longer intergenic regions in the remaining part of the genome. Difference in sizes between the pIIIs indicates that fXo and Xf, which has a predicted pre-pIII of 488 aa (51,036 Da), are distinct phages.
Keywords: Adsorption protein; Aspartic acid and histidine; Gene III; High content of glycine; Membrane-anchoring domain; Signal sequence; Xanthomonas.
Introduction
Several members of Xanthomonas, a genus of gram-negative phytopathogenic bacteria, are known to carry filamentous phages, e.g., fLf, fXv and Cf specifically infecting X. campestris, X. vesicatoria and X. citri, respectively, and Xf and fXo both infecting X. oryzae (Dai et al., 1980; Kuo et al., 1969; Lin et al., 1994; Tseng et al., 1990). Among them, all isolated in Taiwan, the nucleotide sequence has been determined for the fLf and Cf genomes (Kuo et al., 1991; Wen, 1992), whereas the amino acid sequence has been described in an abstract for the Xf gene III coding for the adsorption protein (pIII) (Yang and Yang, 1998), although the sequence was not yet available in the
database. Like other filamentous phages, they possess a circular single-stranded DNA (ssDNA) genome, produce replicative form (RF) during DNA replication, and propagate without lysis of the host cells (Dai et al., 1980; Kuo et al., 1969; Lin et al., 1994; Model and Russel, 1988; Tseng et al., 1990). Several interesting properties of these Xanthomonas phages have also been noticed, including: i) they hold restrictive host specificity, each phage being able to infect only its own host; however, they can propagate in the non-host Xanthomonas cells upon electroporation with RF or ssDNA, and the electroporated cells are capable of releasing authentic phage particles (Lin et al., 1994; Yang and Yang, 1997) indicating that host specificity is determined by the early steps of infection, i.e., adsorption and/or penetration, and ii) the genomes of Cf (7.8 kb), Xf (7.4 kb), and fXo (7.6 kb) are similar in size, but substantially larger than that of fLf (6.0 kb) and fXv (6.4 kb) (Kuo et al., 1991; Lin et al., 1994; Wen, 1992; Yang and Yang, 1998). Although Xf and fXo infect the same host, comparative study of these two phages has not been performed.
In filamentous phages, such as the best studied Ff phages (the closely related M13, f1 and fd), a phage particle contains about 2,700 copies of the major coat protein (pVIII) and three to five copies each of the four minor coat proteins (pIII, pVI, pVII and pIX), with pIII and pVI located at one end and pVII and pIX located at the other
1Present address: Food Industry Research & Development Institute, Hsinchu 300, Taiwan.
2Present address: Pesticide Toxicology Department, Taiwan Agricultural Chemicals and Toxic Substances Research Institute, Wu-Feng, Taichung 413, Taiwan.
3Present address: Center for Biophysics and Computational Biology, University of Illinois at Urbana-Champaign, Urbana, IL 61801, U.S.A.
4Present address: Department of Microbiology, Tzu-Chi University, Hualien 970, Taiwan.
*Corresponding author. Tel: 886-4-285-1885; Fax: 886-4-287-4879; E-mail: yhtseng@dragon.nchu.edu.tw