Bot. Bull. Acad. Sin. (2001) 42: 201-206
Cheng et al. Genetic relationship among peach cultivars
Genetic diversity and relationship among peach cultivars based on Random Amplified Microsatellite Polymorphism (RAMP)
Hsiu-Yu Cheng, Wei-Chen Yang, and Ju-Ying Hsiao1
Deptartment of Botany, National Chung Hsing University, Taichung, Taiwan, Republic of China
(Received August 24, 2000; Accepted January 11, 2001)
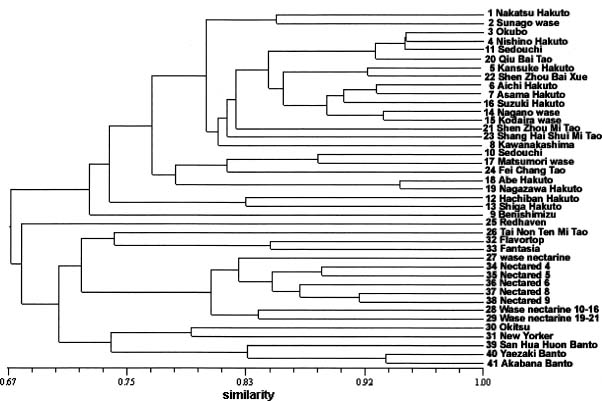
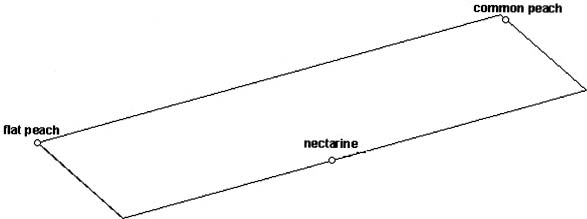
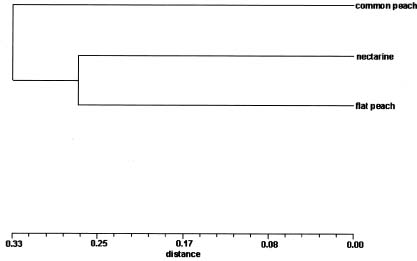
Abstract. Peach (Prunus persica (L.) Batsch) is a common fruit tree species, mainly in the temperate region. It is an important economic plant in Taiwan. In the present study, the genetic relationships among 26 cultivars of common peach (P. persica var. vulgaris Maxim.), 12 cultivars of nectarine (P. persica var. nectarina Maxim.), and three cultivars of flat peach (P. persica var. platycarpa Bailey) were estimated using RAMP markers. Eighty-two polymorphic bands were obtained using ten combinations of primers. The cluster analysis based on RAMP data revealed that the groupings were generally consistent with the classification of the varieties and the regions of origin of cultivars. The common peach cultivars originating in China and Japan formed a cluster. A possible explanation is that the Japanese cultivars may be developed from cultivars introduced from China. Within flat peach cultivars, the groupings also indicated that the genetic relationship among cultivars is correlated with the regions of origin of cultivars. Analysis of molecular variance (AMOVA) revealed that the variance components among and within three peach groups expressed as percentages of the total variation were 30.3% and 69.7%, respectively.
Keywords: Peach; Prunus persica; Genetic relationship; RAMP; AMOVA.
Introduction
Peach (Prunus persica (L.) Batsch) is a common fruit tree in temperate regions. It is considered the queen of temperate-zone fruits and, next to apple, is the world's most widely grown fruit tree (Bailey and Bailey, 1976). Three varieties can be recognized taxonomically based on fruit morphology. The common peach (P. persica var. vulgaris Maxim.) has rounded and hairy fruits. The nectarine (P. persica var. nectarina Maxim.) has rounded fruits without hairs. The flat peach (P. persica var. platycarpa Bailey) has flat fruits. Peach is one of the important economic plants in Taiwan. Besides traditional cultivars, many cultivars of common peach and nectarine were introduced from Japan and United States recently and grown in high altitude areas. The over-utilization of high altitude mountain slopes resulted in soil erosion and the destruction of natural environments. Peach breeding programs in Taiwan, aimed at producing cultivars suitable for lower altitudes, could reduce the destruction of natural environments. Molecular markers such as isozymes (Arulsekar et al., 1986; Durham et al., 1987; Chaparro et al., 1994), RFLP (Eldredge et al., 1992; Rajapakse et al., 1995; Quarta et al., 1996), and RAPD (Chaparro et al., 1994; Rajapakse et al., 1995; Warburton et al., 1996), have been useful in estimating the genetic relationship and in the genetic linkage mapping of the peach genome. The estimation of genetic relationships among cultivars provides the basic information for
breeding programs. Molecular markers could also be used in assisting the process of artificial selection. Random amplified microsatellite polymorphism (RAMP; Wu et al., 1994) has been demonstrated to be another potentially valuable molecular marker for the study of genetic relationships in cultivated plant species. The combination of a simple sequence repeat (SSR; microsatellite) and a random sequence was used to amplify genomic DNA fragments in RAMP. RAMP has been employed in studies of the cultivars of barley (Wu et al., 1994; Becker and Heun, 1995; Sanchez de la Hoz et al., 1996). The usefulness of the RAMP molecular marker has not been widely tested in other plant species. The objectives of the present study are to investigate the genetic relationship of some peach cultivars available in Taiwan and to estimate the genetic diversity among and within major peach groups using RAMP markers.
Materials and Methods
The leaf samples of 41 peach cultivars (Table 1) were collected from the Lona division of the Taiwan Agriculture Research Institute and Mountain Experimental Farm of National Taiwan University and stored at -70°C before DNA extraction. DNA extraction followed the method of Doyle and Doyle (1990). The DNA concentration was measured using a Hoefer TKO 100 fluorometer with Hoechst dye solution. Ten combinations of primers selected between three SSR-primers (RM21, RM23, RM24) and five random primers (A1, A4, B1, Q5, V6; Operon Tech. Inc., USA) were used for RAMP amplifications. The five random primers were also used singularly for RAPD amplifi
1Corresponding author. Tel: 886-4-22840417 ext. 315; Fax: 886-4-22874740; E-mail: jyhsiao@dragon.nchu.edu.tw