Bot. Bull. Acad. Sin. (2001) 42: 251-258
El-Shora Properties and immobilization of urease
Properties and immobilization of urease from leaves of Chenopodium album (C3)
H.M. El-Shora
Botany Department, Faculty of Science, Mansoura University, Mansoura, Egypt
(Received October 11, 2000; Accepted April 16, 2001)
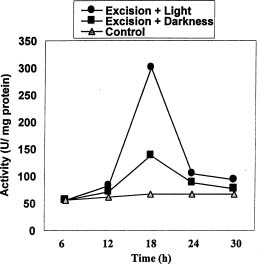
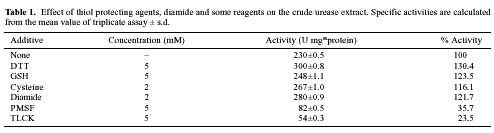
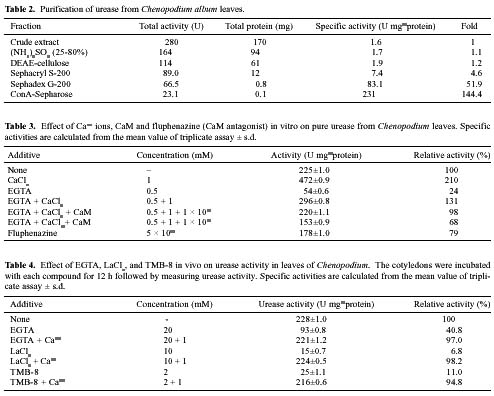
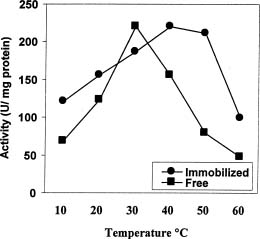
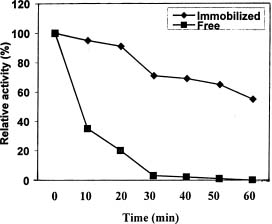
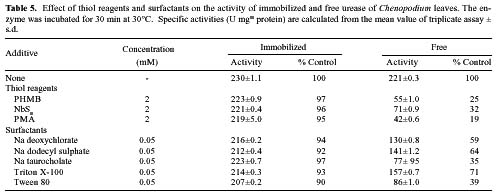
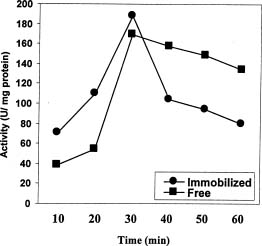
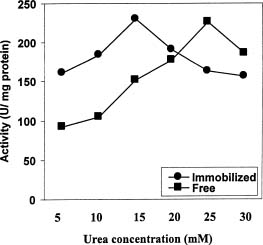
Abstract. In Chenopodium album, leaf excision and light both increase urease (EC. 3.5.1.5) activity. Dithiothreitol (DTT), reduced glutathione (GSH), cysteine and diazoinedicarboxylic acid bis(N,N-dimethylamide) (diamide) activated the crude enzyme. In contrast, crude urease was inhibited by phenylmethylsulphonyl fluoride (PMSF) and N-a-p -tosyl-L-lysine chloromethyl ketone HCl (TLCK), suggesting the presence of serine and histidine residues in the active site. The enzyme is Ca dependent for its activity and exogenous calmodulin (CaM) did not stimulate it. However the enzyme is strongly inhibited by CaM antagonist fluphenazine, indicating the presence of a Ca-like domain. EGTA, LaCl3 and tetraacetic acid, 3,4,5,-trimethoxybenzoic acid 8-(diethyl-amino)-octyl ester (TMB-8) inhibited urease activity in vivo, and the inhibition was restored by exogenous Ca. Urease was immobilized in gelatin by covalent cross-linking with formaldehyde as organic hardener. The results indicated enhanced resistance to thermal denaturation, increased temperature optima (from 30°C to 40°C), and a rapid rate of substrate saturation were achieved after immobilization. The free urease showed remarkable loss of activity in the presence of sodium dodecyl sulphate, sodium deoxycholate, sodium taurocholate, Triton X-100, and Tween 80. The free enzyme lost 68%, 75% and 81% of its activity in the presence of 5,5´-dithiobis-(2-nitrobenzoic acid) (NBS2), p-hydroxymercuribenzoate (PHMB) and phenylmercuric acetate (PMA) as thiol reagents. However, the immobilized enzyme was not affected significantly by these compounds. By increasing the incubation time, the activity of immobilized enzyme decreased faster than that of the free one.
Keywords: Chenopodium album; Immobilization; Properties; Purification; Urease.
Abbreviations: DTT, dithiothreitol; GSH, reduced glutathione; diamide, diazoinedicarboxylic acid bis(N,N-dimethylamide); PMSF, phenylmethylsulphonyl fluoride; TLCK, N-a-p -tosyl-L-lysine chloromethyl ketone HCl; TMB-8, tetraacetic acid, 3,4,5,-trimethoxybenzoic acid 8-(diethyl-amino)-octyl ester; PCM, phenylmercuric acetate; NBS2, 5,5´-dithiobis-(2-nitrobenzoic acid); PHMB, p-hydroxymercuribenzoate; SDS-PAGE, sodium dodecyl sulphate polyacrylamide gel electrophoresis.
Introduction
It has been proposed that plant urease functions in the assimilation of urea, normally formed in plants as a result of the hydrolysis of arginine to ornithine catalyzed by the enzyme arginase (Polacco and Holland, 1993; Lea, 1997). The enzyme catalyses the hydrolysis of urea to ammonia and carbon dioxide.
The enzyme has been found in the leaves, roots, and bark of plants with actively growing tissues possessing greater activity than senescing ones (Thompson, 1980; Horgan et al., 1983). The enzyme activity in the leaf is also important if foliar application of urea is considered as fertilizer. It has been proposed that foliar treatment with urea increases total leaf yield in mulberry plants, improves leaf nutritional quality, and leads to an increase of cocoon yield (Fotedar and Chakrabarty, 1985; Sarker and Absar, 1995). Nevertheless, the information available on properties of plant leaf urease is quite incomplete.
Genetic and chemical blocking of leaf urease activity caused necrotic leaf tip, associated with urea accumulation (Eskew et al., 1983; Krogmeier et al., 1989; Stebbins et al., 1991). This reveals that urease plays a significant role in urea metabolism.
Interestingly, urease has been partially purified from soybean leaf (Kerr et al., 1983; Polacco and Winkler, 1984; Hirayama et al., 2000) and seeds of several leguminous plants (Reithel, 1971; Polacco and Havir, 1979). However, the leaf urease yielded was biochemically distinct from that of seed enzyme (Polacco and Winkler, 1984).
Urease from jack bean seed, soybean seeds, and bacteria have a lower optimum pH, at pH 7-8 (Kerr et al., 1983; Mobley and Hausinger, 1989), whereas that of most fungal urease ranges between 8-8.5 (Mobley et al., 1995; Lubbers et al., 1996). Interestingly, urease partially purified from soybean leaves had two pH optima (5.3 and 8.8) (Kerr et al., 1983).
There have been few reports on the purification of urease to homogeneity from a plant leaf source despite its significant role in urea metabolism. This is likely owing
Fax: 050/346781 (Mansoura); E-mail: Sinfac@mum.mans.eun.eg