Bot. Bull. Acad. Sin. (2001) 42: 265-272
Chang and Yu Ozone resistance of bedding plants
Correlation between ozone resistance and relative chlorophyll fluorescence or relative stomatal conductance of bedding plants
Yu-Sen Chang1,* and M.R. Yu
Department of Horticulture, National Taiwan University, No. 1, Section 4. Roosevelt Road, Taipei 10617, Taiwan, Republic of China
(Received December 11, 2000; Accepted July 20, 2001)
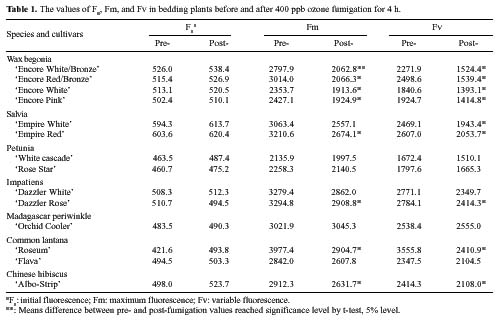
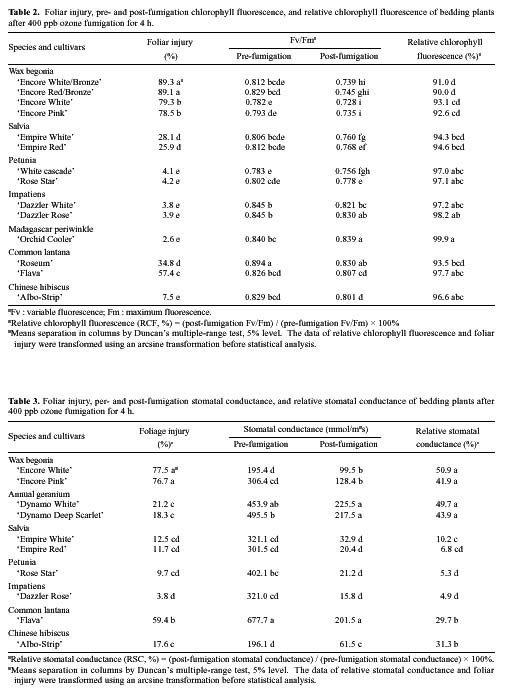
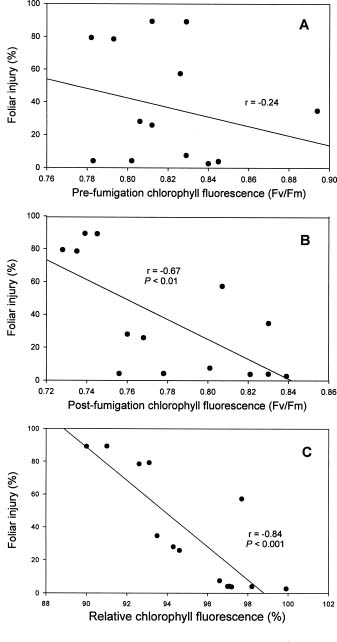
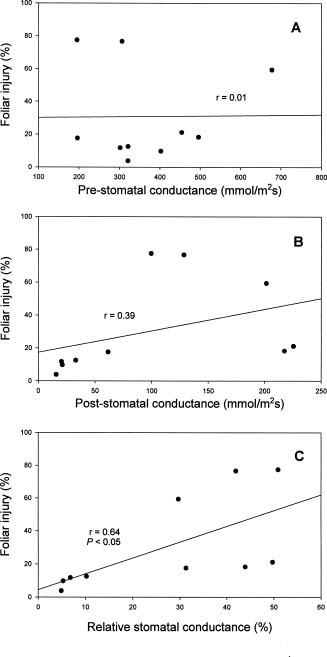
Abstract. Eight species of bedding plants were exposed to 400 ppb ozone (O3) for 4 h. On the basis of the resulting foliar injury, Madagascar periwinkle (Catharanthus roseus) and impatiens (Impatiens walleriana) were the most resistant to O3, and wax begonia (Begonia×semperflorens-cultorum) was the most sensitive to O3. Clorophyll fluorescence (Fv/Fm) and stomatal conductance were measured before and after the O3 fumigation. There was a significant regression between, the degree of foliage injury by O3 and relative chlorophyll fluorescence (RCF, ratio of post-fumigation Fv/Fm: pre-fumigation Fv/Fm) or relative stomatal conductance (RSC, ratio of post-fumigation stomatal conductance: pre-fumigation stomatal conductance) (r = - 0.84, P<0.001 and r=0.64, P<0.05 respectively). That is, species of tested bedding plants that had stronger O3 resistance generally had higher RCF and lower RSC values. It is suggested that RCF and RSC measurements could serve as indicators to screen plants for O3 resistance.
Keywords: Air pollution; Bedding plants; Chlorophyll fluorescence; Ozone resistance; Stomatal conductance.
Introduction
Ozone (O3) is an important and widespread phytotoxic air pollutant (Heck et al., 1986; Ormrod and Hale, 1995). It can reduce the growth rate of plants and lower crop yield and induce visible foliage injury (Heagle, 1989). Photosynthesis is a core function in the physiology of all plants and is certainly a primary target of O3 effects even if it is not clear what mechanisms are involved in the limitation of this process (Heath, 1994). Measurements of photosynthesis have therefore often been used in the assessment of O3 injury (Heath, 1996). As part of the methodology assessed for the study of photosynthetic process, chlorophyll fluorescence represents a useful and non-destructive tool for in vivo stress detection (Owens, 1994), and it is widely used to study the effects of O3 on the photosynthetic process, especially photosystem 2 (PS2) in light reaction (Schreiber et al., 1978; Lee, 1991; Guidi et al., 1993, 1997).
Vegetation responses to O3 are dependent on both uptake or flux of O3 into the leaf and the action of defensive mechanisms in plant tissue. Defensive mechanisms operating within plant tissue to detoxify O3 or repair injured tissue are an important component of plant O3 resistance, but they are complex and difficult to quantify (Musselman
and Massman, 1999). O3 generally leads to varying degrees of stomatal closure and reduces stomatal conductance (Lehnherr et al., 1987; Mansfield and Pearson, 1996; Guidi et al., 1997). Aben et al. (1990) have shown direct effects of O3 on stomata as well as on photosynthesis, but stomata appeared to be more sensitive. In addition, O3 enters plant tissue primarily through the stomata, so the first step toward control of O3 injury depends on stomatal conductance (Ormrod and Hale, 1995). The change of stomatal conductance may be regarded as a kind of resistance mechanism (Mansfield and Freer-Smith, 1984; Reiling and Davison, 1995). Since standard techniques are available to quantify stomatal conductance, monitoring stomatal behaviour may be a possible method of proving variations in O3 resistance.
The objectives of this study were to: (1) screen O3 resistance in terms of foliar injury in tested bedding plants; (2) study the correlation between foliar injury and the parameters of chlorophyll fluorescence or stomatal conductance; (3) evaluate the feasibility of using the parameters of chlorophyll fluorescence or stomatal conductance as indicators of O3 resistance in bedding plants.
Materials and Methods
Relationship between Ozone Resistance and Chlorophyll Fluorescence of Bedding Plants
Plant materials. Plant species used in this study were wax begonia (Begonia×semperflorens-cultorum) `Encore Red/Bronze', `Encore Red/Bronze', `Encore White', and
1Associate Professor and Graduate student of the Department of Horticulture, National Taiwan University.
*Corresponding author. Tel: 866-2-23630231-3340; Fax: 866-2-23635849; E-mail: yschang@ccms.ntu.edu.tw