Bot. Bull. Acad. Sin. (2001) 42: 287-302
Yang et al. Coccolithophorid and coccolith distribution
Distribution of coccolithophorids and coccoliths in surface ocean off northeastern Taiwan
Tien-Nan Yang1,3, Kuo-Yen Wei1,*, and Gwo-Ching Gong2
1Global Change Research Center and Department of Geosciences, National Taiwan University, Taipei, 10716 Taiwan, ROC
2Department of Oceanography, National Taiwan Ocean University, Keelung, Taiwan, ROC
(Received October 27, 2000; Accepted July 26, 2001)
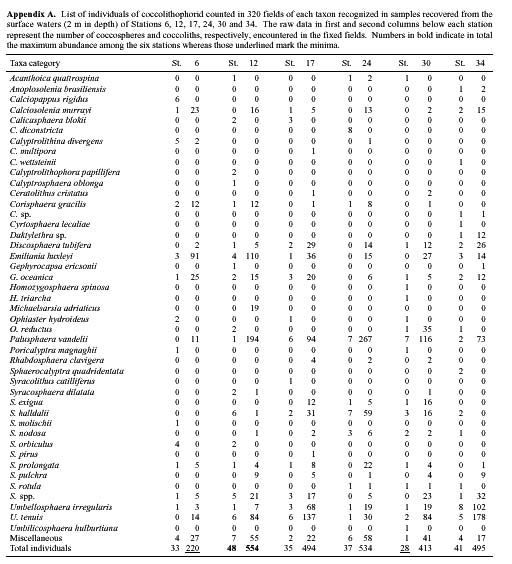
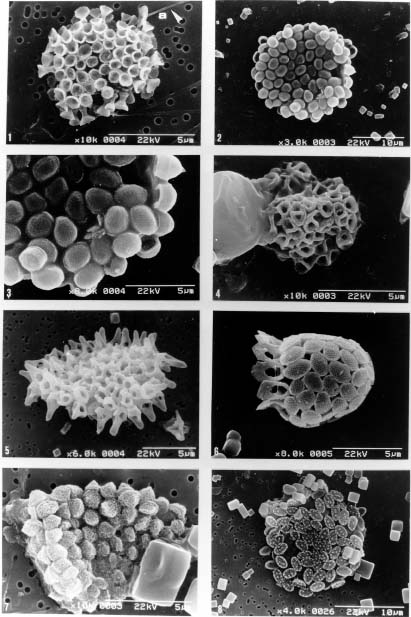
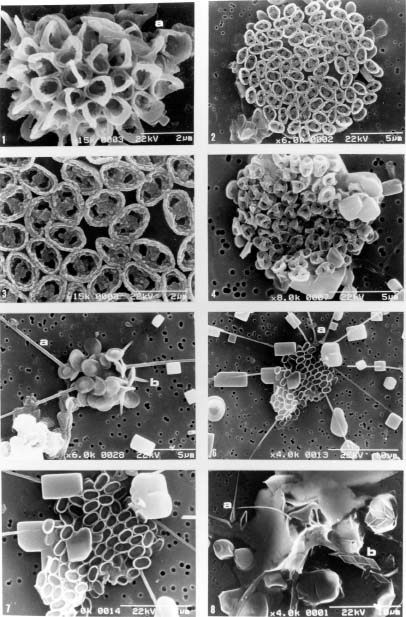
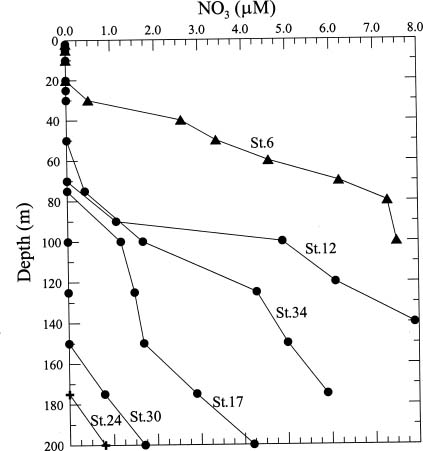
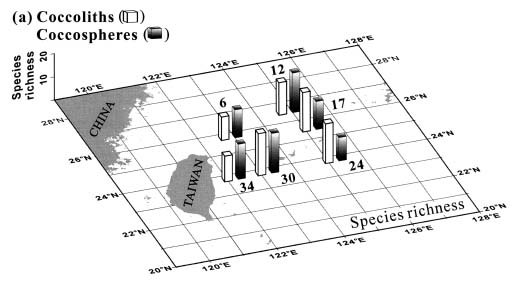
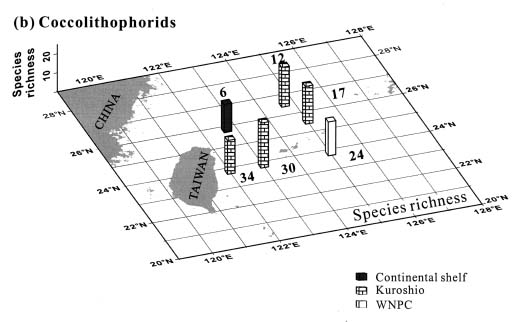
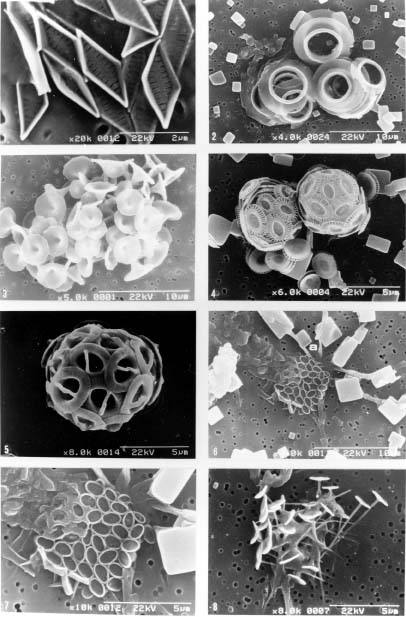
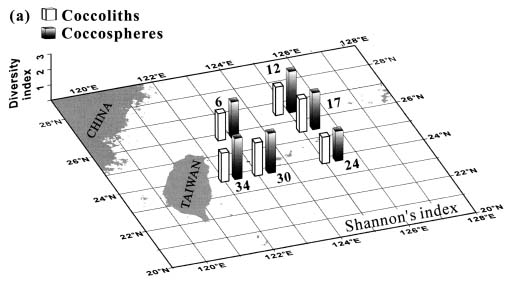
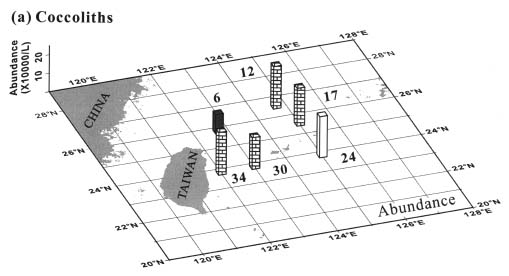
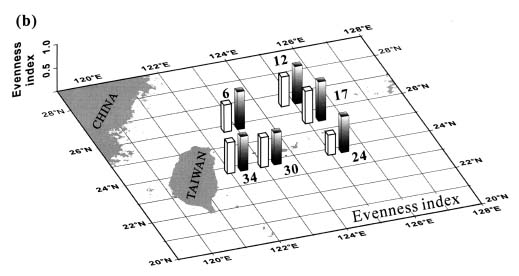
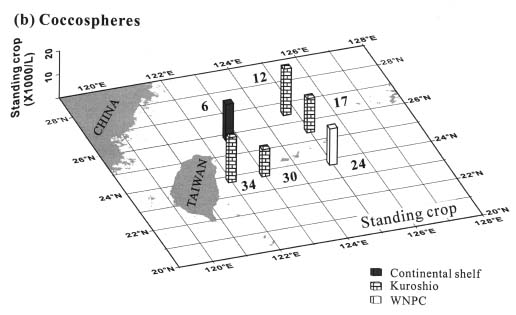
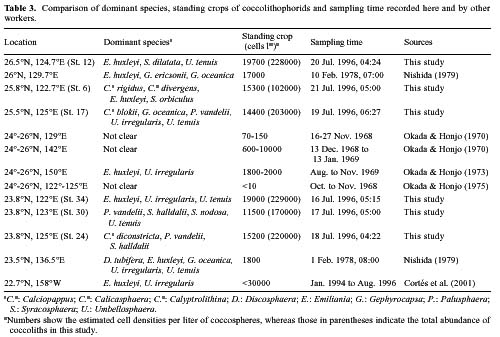
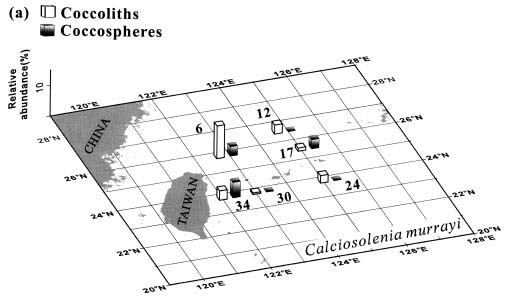
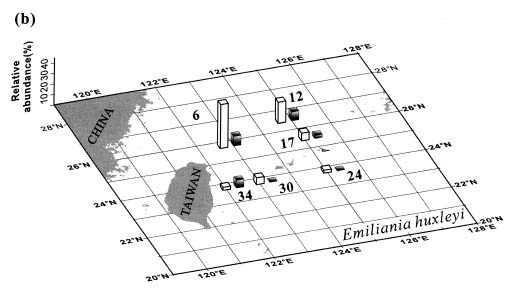
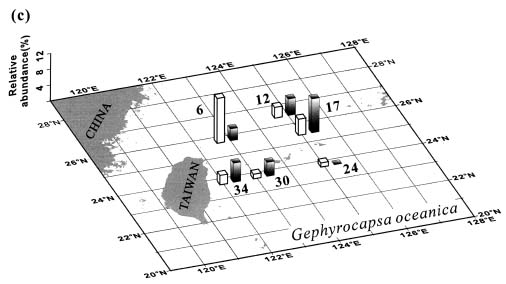
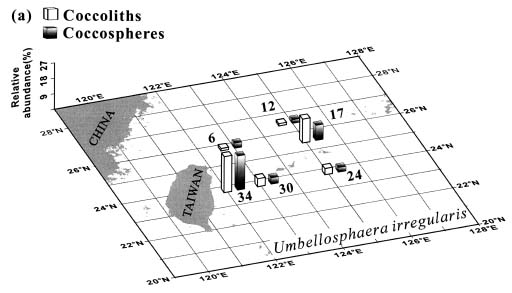
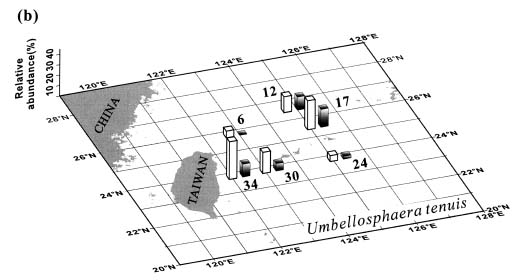
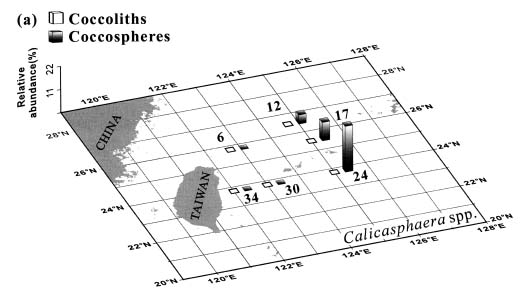
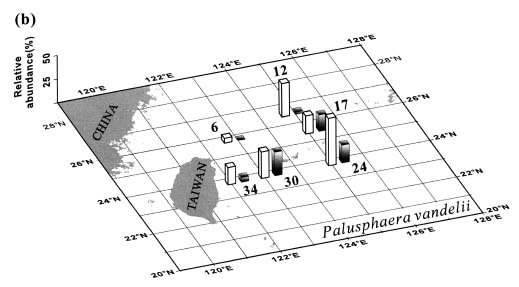
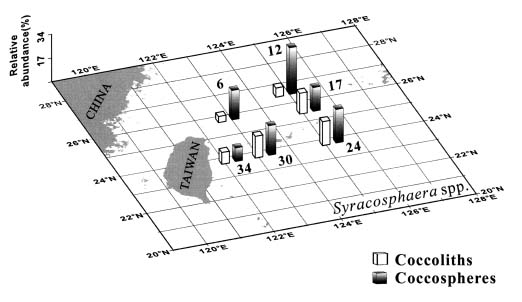
Abstract. This study depicts quantitatively the distribution of coccolithophorids and coccoliths during one summer season in the area off northeastern Taiwan where the Kuroshio flows northward and interacts with the shelf waters of the East China Sea. To minimize the influence of diurnal variation of coccolithophorids, only samples taken at sunrise were analyzed. Sea-surface water (2 m in depth) samples were obtained at six stations during the summer of 1996. Forty-one species were identified with Emiliania huxleyi (Lohmann) Hay et Mohler, Palusphaera vandelii Lecal emend. R. E. Norris, Umbellosphaera Paasche spp. and Syracosphaera Lohmann spp. being the predominant forms. Three coccolithophorid communities were recognized: (1) the continental shelf community, dominated by Emiliania huxleyi, Gephyrocapsa oceanica Kamptner and Calciosolenia murrayi Gran, which showed intermediate biodiversity and species evenness; (2) the Kuroshio community, which showed the highest diversity and evenness, with a flora dominated by genus Umbellosphaera Paasche; and (3) the Western North Pacific Central Water community, which had the lowest diversity and evenness, with dominant species Calicasphaera Kleijne and Palusphaera vandelii. The absolute abundance of loose coccoliths ranged from 10.2 × 104 individual coccoliths l-1 to 22.9 × 104 individual coccoliths l-1, while those of coccospheres were much less, ranging from 11.5 × 103 cells l-1 to 19.7 × 103 cells l-1. The largest absolute abundance of coccoliths and coccospheres was found in the Kuroshio path.
Keywords: Coccolithophorid; Diversity; East China Sea; Kuroshio; Standing crops.
Introduction
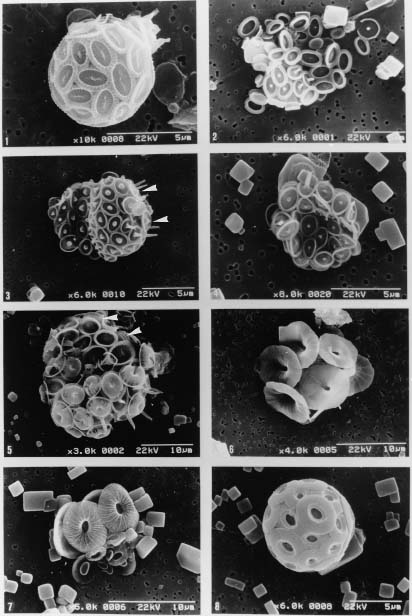
Coccolithophorids are unicellular, marine, golden-brown algae (Haptophyta) commonly found in near-surface waters in patchy distributions. They are one of the world's major primary producers, contributing about 15 per cent of the average oceanic phytoplankton biomass to the oceans (Berger, 1976). They also produce elaborate, minute calcite platelets (coccoliths), covering the cell to form a coccosphere and supplying up to 60 per cent of the bulk pelagic calcite deposited on the sea floors (Honjo, 1996). Coccoliths can reflect the visible light from surface water during coccolithophorid bloom season (Holligan et al., 1983; Groom and Holligan, 1987; Balch et al., 1991) and thus enhance albedo. Coccolithophorids, therefore, have recently gained much attention as important players in global climate change and carbon cycles (Westbroek et al., 1993 and references therein; Hagino et al., 2000; Cortés et al., 2001; Haidar and Thierstein, 2001).
The large-scale distribution of modern coccolithophorids in the open ocean has been studied in various
parts of the world (for review, see Winter et al., 1994 and references therein). About two decades ago, Okada and Honjo (1970; 1973) and Nishida (1979) completed large-scale studies concerning coccolithophorid biogeography in the Pacific Ocean, with emphasis on the North Pacific. Okada and Honjo (1973) established six coccolithophorid zones in the surface water along the 155°W meridian based on the distribution pattern of characteristic species. Recently, Hagino et al. (2000) documented the spatial dynamics of coccolithophorid assemblages in the Western-Central Equatorial Pacific Ocean, while Cortés et al. (2001) studied the temporal variation pattern of coccolithophorid ecology in water column at the HOT station ALOHA, Hawaii. For the western boundary constraint of the Pacific, Okada and Honjo (1975) conducted a large-scale study of coccolithophorid biogeography of the western Pacific marginal seas. Nevertheless, few detailed analyses of living coccolithophorids of individual marginal seas and biogeographic provinces of the western Pacific have appeared. The present study focuses particularly on the community structure of coccolithophorids in the surface waters (~2 m in depth, hereafter defined as the surface water) of summer-season in the area offshore of northeastern Taiwan, where the northward flowing Kuroshio encounters the Ryukyu Ridge, collides with the continental shelf break, and then deflects to the northeast (Nitani, 1972).
3E-mail: f4224103@ms.cc.ntu.edu.tw
*Corresponding author. Tel: +886-2-23691143; Fax: +886-2-23636095; E-mail: weiky@ms.cc.ntu.edu.tw