Bot. Bull. Acad. Sin. (2002) 43: 21-29
Liou et al. DNA fingerprint analysis of Phytophthora parasitica
Analysis of Phytophthora parasitica by retrotransposon-derived DNA fingerprinting
Ruey-Fen Liou1,*, Jent-Turn Lee1, Hsiao-Ching Lee1, and Pao-Jen Ann2
1Department of Plant Pathology, National Taiwan University, Taipei 106, Taiwan
2Department of Plant Pathology, Taiwan Agricultural Research Institute, Wu Fong 413, Taiwan
(Received August 2, 2001; Accepted September 14, 2001)
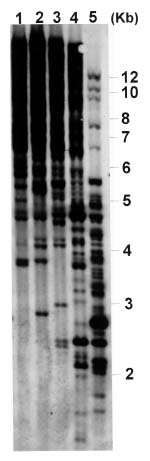
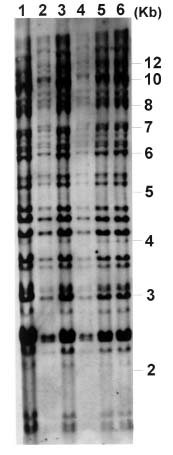
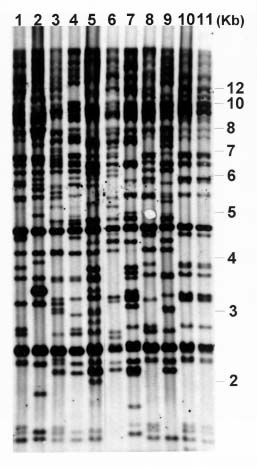
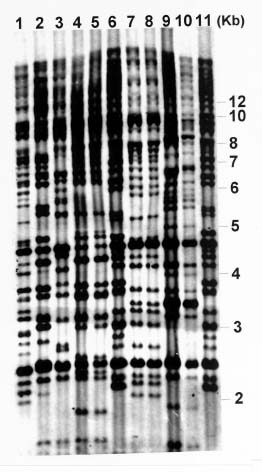
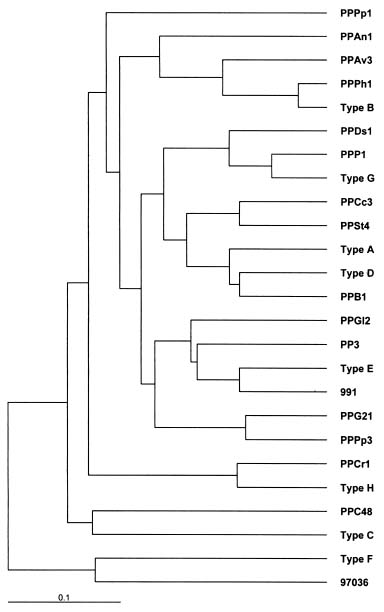
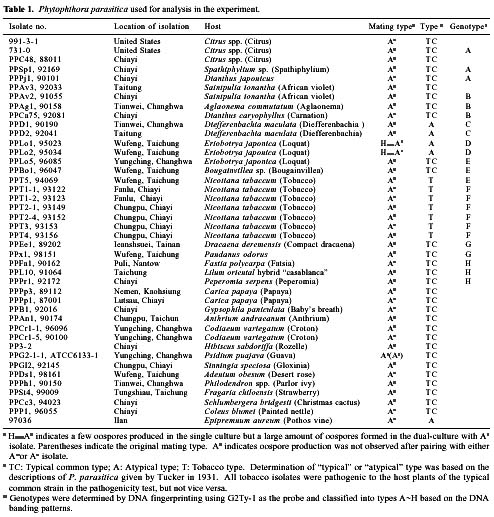
Abstract. Phytophthora parasitica, being able to attack a wide variety of plants, is a very important plant pathogen in Taiwan. It has been shown that isolates of P. parasitica from tobacco, dieffenbachia, and loquat differed significantly from other isolates in morphology and pathogenicity and were recognized as "atypical" types of the fungus. In the current study, a partial sequence of the reverse transcriptase gene of a Ty1-copia retrotransposon was cloned from P. parasitica and used as the probe, designated herein as G2Ty-1, for genomic Southern hybridization analyses of P. parasitica. It was demonstrated that G2Ty-1 existed as a moderately repetitive sequence in the genome of P. parasitica. The banding pattern of G2Ty-1 was mitotically stable for at least five consecutive asexual generations. DNA fingerprint analysis of P. parasitica using G2Ty-1 as the probe demonstrated that the banding patterns of G2Ty-1 were highly polymorphic among the isolates analyzed. "Atypical" strains from tobacco, dieffenbachia, and loquat, nonetheless, displayed host-specific banding patterns. Phylogram generated by cluster analysis demonstrated that isolates from tobacco and dieffenbachia were well separated from each other and from all other isolates of P. parasitica. These results provided a genetic basis for distinguishing these isolates from others and validate the use of retrotransposon as a genetic marker to study phylogeny in Phytophthora parasitica.
Keywords: DNA fingerprinting; Genotypes; Host specificity, Phytophthora parasitica; Retrotransposon.
Introduction
Phytophthora parasitica Dastur (= Phytophthora nicotianae Breda de Haan), an oomyceteous fungus, is the most widespread and important species of Phytophthora in Taiwan (Ho et al., 1995). It causes root rot, foot rot, leaf blight, and fruit rot in a variety of economically important crops (Ann et al., 1992; Ann, 1992, 2000a, 2000b; Erwin and Ribeiro, 1996). The taxonomy of Phytophthora is based mainly on morphological and physiological characteristics (Waterhouse, 1963). Although isolates morphologically identified as P. parasitica are pathogenic to a wide range of plant species, considerable evidence shows that some isolates are specifically pathogenic to a few hosts or a single host, such as tobacco (Bonnet et al., 1978, 1994; Matheron and Matejka, 1990; Erwin and Ribeiro, 1996). In support of this view, it was demonstrated in a recent study that isolates from dieffenbachia (Dieffenbachia maculata Don) were nonpathogenic or weakly pathogenic to other hosts of P. parasitica, nor were isolates from these plants pathogenic to dieffenbachia (Ann, 1992). Besides, some isolates from loquat (Eriobotrya japonica (Thunb.) Lindl.), known as "atypical" strains of P. parasitica, were found to be highly
pathogenic to loquat while displaying reduced virulence toward eggplant fruits, sweet orange, and tomato seedlings as compared to typical strains in the pathogenicity test (Chern et al., 1998). It is not clear if atypical characteristics in pathogenicity observed in these isolates result from differentiation of the fungi at the genetic level. In past years, molecular markers such as isozyme patterns (Oudemans and Coffey, 1991) or mitochondrial restriction fragment length polymorphism (RFLP) (Föster et al., 1990; Lacourt et al., 1994) have been used for analysis of P. parasitica. These studies, however, revealed only a low level of polymorphism among the P. parasitica isolates analyzed, and no correlation with pathogenicity was observed. As a first attempt to seek molecular support for host specialization in P. parasitica isolates, retrotransposon-derived DNA fingerprinting was used to analyze the genotypes of various P. parasitica isolates in the current study.
Retrotransposons are mobile genetic elements, which replicate through an RNA intermediate prior to integration into the chromosomal DNA of their host. As a result, they may constitute a substantial portion of the repetitive sequences in an organism, and provide useful markers for analyses of the plant and fungal genomes (Hamer et al., 1989; Purugganan and Wessler, 1995; Flavell et al., 1998). According to the structural organization and the amino acid sequences of the encoded reverse transcriptase and
*Corresponding author. Fax: 886-2-23620271; E-mail: rfliou@ccms.ntu.edu.tw