Bot. Bull. Acad. Sin. (2002) 43: 63-68
Hao et al. Taxonomical relationships in Allium sect. Sacculiferum
A study of taxonomical relationships among species of Korean Allium sect. Sacculiferum (Alliaceae) and related species using inter-simple sequence repeat (ISSR) markers
Gang Hao1, Dong-Hee Lee2, June Seung Lee2, and Nam Sook Lee2,*
1South China Institute of Botany, The Chinese Academy of Sciences, Guangzhou 510650, P. R. China
2Department of Biology, Ewha Womans University, Seoul 120-750, Korea
(Received July 2, 2001; Accepted September 13, 2001)
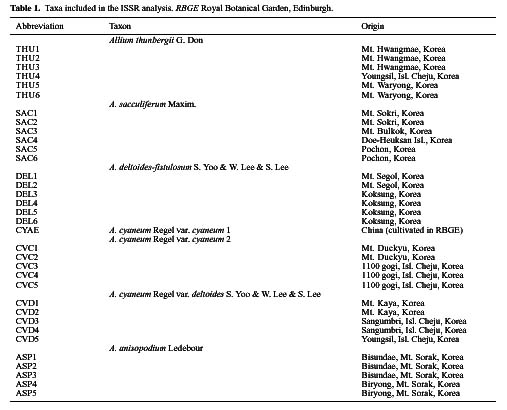
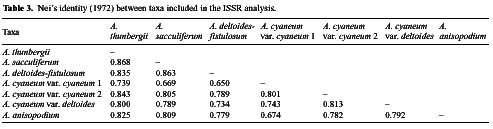
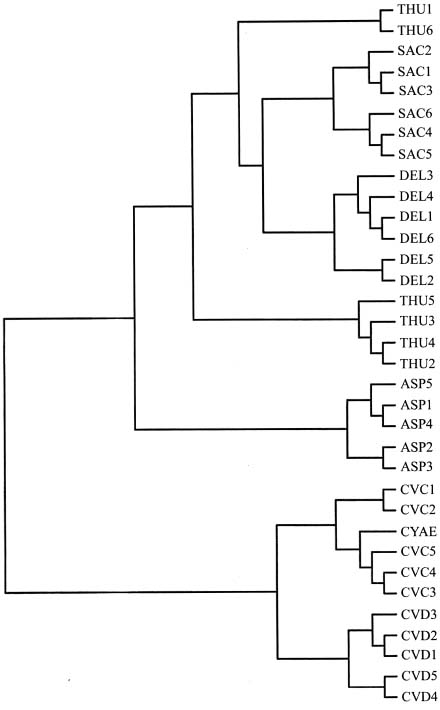
Abstract. Morphological differences among Korean Allium subgenus Rhizirideum sect. Sacculiferum and some related species are minor, making species delimitation difficult. Inter-simple sequence repeat (ISSR) was employed to assess genetic diversity and relationships. Thirty-four accessions, representing six taxa of A. thunbergii, A. sacculiferum, A. deltoides-fistulosum, A. cyaneum var. cyaneum, A. cyaneum var. deltoides, A. anisopodium, were sampled in this study. The ISSR markers revealed high polymorphism among taxa studied. In the phenetic analysis, accessions of A. sacculiferum and A. deltoides-fistulosum are nested within A. thunbergii accession group; A. cyaneum var. cyaneum and A. cyaneum var. deltoides are separated from the above sect. Sacculiferum group; the status of putative A. anisopodium is less clear, with a position between sect. Sacculiferum and A. cyaneum. We propose that A. deltoides-fistulosum and A. sacculiferum be sunk into A. thunbergii and that the putative A. anisopodium be recognized as A. cyaneum var. deltoides.
Keywords: Allium; Genetic diversity; Korea; Phenetic analysis; Sect. Sacculiferum.
Introduction
Section Sacculiferum is small in the large subgenus Rhizirideum of Allium, but poorly delimited. When Gritzenko (1979) proposed this section, it included two species, A. sacculiferum Maxim. and A. komarovianum Vved, both characterized by globose to ovate bulbs, simple coriaceous bulb coats, 3-angular or keeled flat leaves, subglobose rose or violet flowers, simple stamens much longer than the tepals, deep nectary grooves at the base of the ovary covered by hood-like projections, and finally flat obovate seeds (Hanelt and Fritsch, 1994). Gritzenko's (1979) two species were subsequently sunk into A. thunbergii, a rather broadly circumscribed species (Xu, 1980; Xu et al., 1990). Xu (1980) erroneously placed A. thunbergii in sect. Haplostemen, together with the commonly cultivated East Asiatic species A. chinense, a very close relative. In the most recent infrageneric classification of Allium, Hanelt and Fritsch (1994) broadened the concept of sect. Sacculiferum by including A. chinense, and A. virgunculae, a related Japanese species, within it.
In the revision of Korean Allium (Yu et al., 1981), eight species and one variety were recognized in subgenus Rhizirideum, viz: A. thunbergii, A. sacculiferum, A. deltoides-fistulosum (sect. Sacculiferum), A. cyaneum var. cyaneum, A. cyaneum var. deltoides, A. splendens (sect.
Reticulato-bulbosa), A. anisopodium (sect. Anisopodia), A. senescens (sect. Rhizirideum), A. victorialis (sect. Anguinum). Morphologically A. splendens, with its tooth-like scale of the inner filament, and the latter two species, A. senescens with stalk before flowering, and A. victorialis having flat ovate shaped leaf, are distinct from others in the Korean taxa of subgen. Rhizirideum. For sect. Sacculiferum, the species status of A. sacculiferum was maintained, and A. deltoides-fistulosum was newly described (Yu et al., 1981). The morphological differences among most of those species, however, are minor, making species identification difficult. For example, some populations occurring in the Mt. Dukyu region were labeled A. cyaneum var. cyaneum with interstamenous appendage and wax powders on the leaf surfaces, and cross section of leaves semi-circular and hollow, while populations in Mt. Kaya and Isl. Cheju were proposed as A. cyaneum var. deltoides without interstamenous appendage and wax-powders on the leaf surfaces, and cross section of leaves triangular and hollow. Populations distributed in Mt. Sorak were recognized as A. anisopodium with leaf cross-section narrow and flat and solid (Yu et al., 1981), despite the fact that they resemble A. cyaneum var. deltoides in many other respects (Woo, 2000).
Molecular markers have proven useful in clarifying genetic relationships within subgenus Rhizirideum (Dubouzet et al., 1997; Raamsdonk et al., 200), but sect. Sacculiferum remains intact in those studies. Until recently, a preliminary phylogenetic study of sect.
*Corresponding author. Tel: 82-2-3277 2366; Fax: 82-2-3277 2385; E-mail: namsook@mm.ewha.ac.kr