Ho et al. Genetic relationships of Casuarina equisetifolia
Bot. Bull. Acad. Sin. (2002) 43: 93-98
An assessment of DNA polymorphisms and genetic relationships of Casuarina equisetifolia using RAPD markers
Kuen-Yih Ho1, Chern-Hsiung Ou2, Jenq-Chuan Yang3, and Ju-Ying Hsiao4,*
1Chung-Pu Station of Taiwan Forestry Research Institute, 15 Shih-Yen Road, Hunshui, Chung-Pu, Chiayi County 606, Taiwan, Republic of China
2Department of Forestry, National Chung Hsing University, Taichung, Taiwan, Republic of China
3Taiwan Forestry Research Institute, 53 Nanhai Road, Taipei 100, Taiwan, Republic of China
4Department of Botany, National Chung Hsing University, Taichung 402, Taiwan, Republic of China
(Received August 3, 2001; Accepted October 29, 2001)
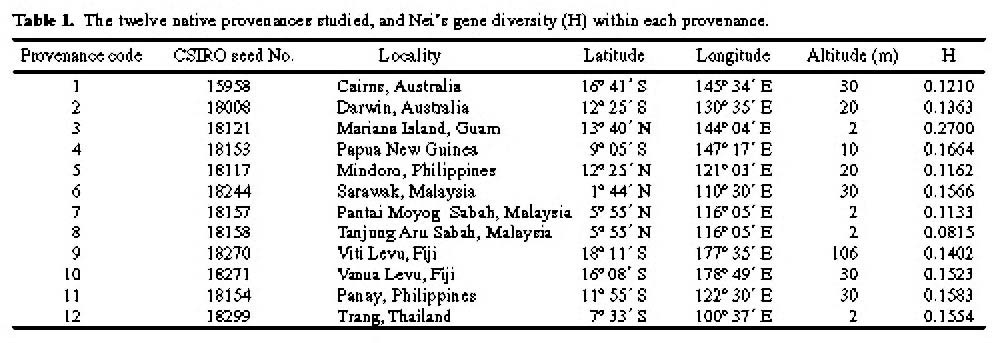
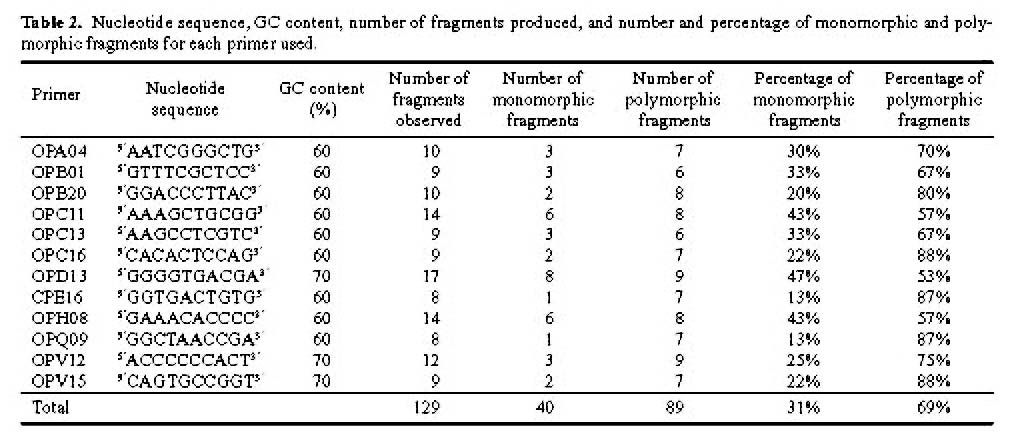
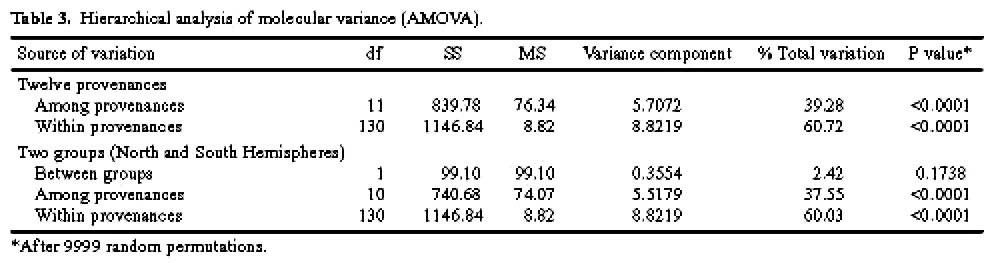
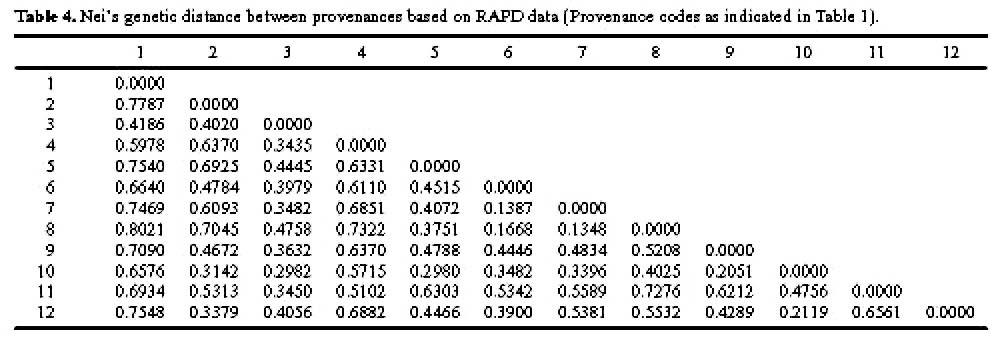
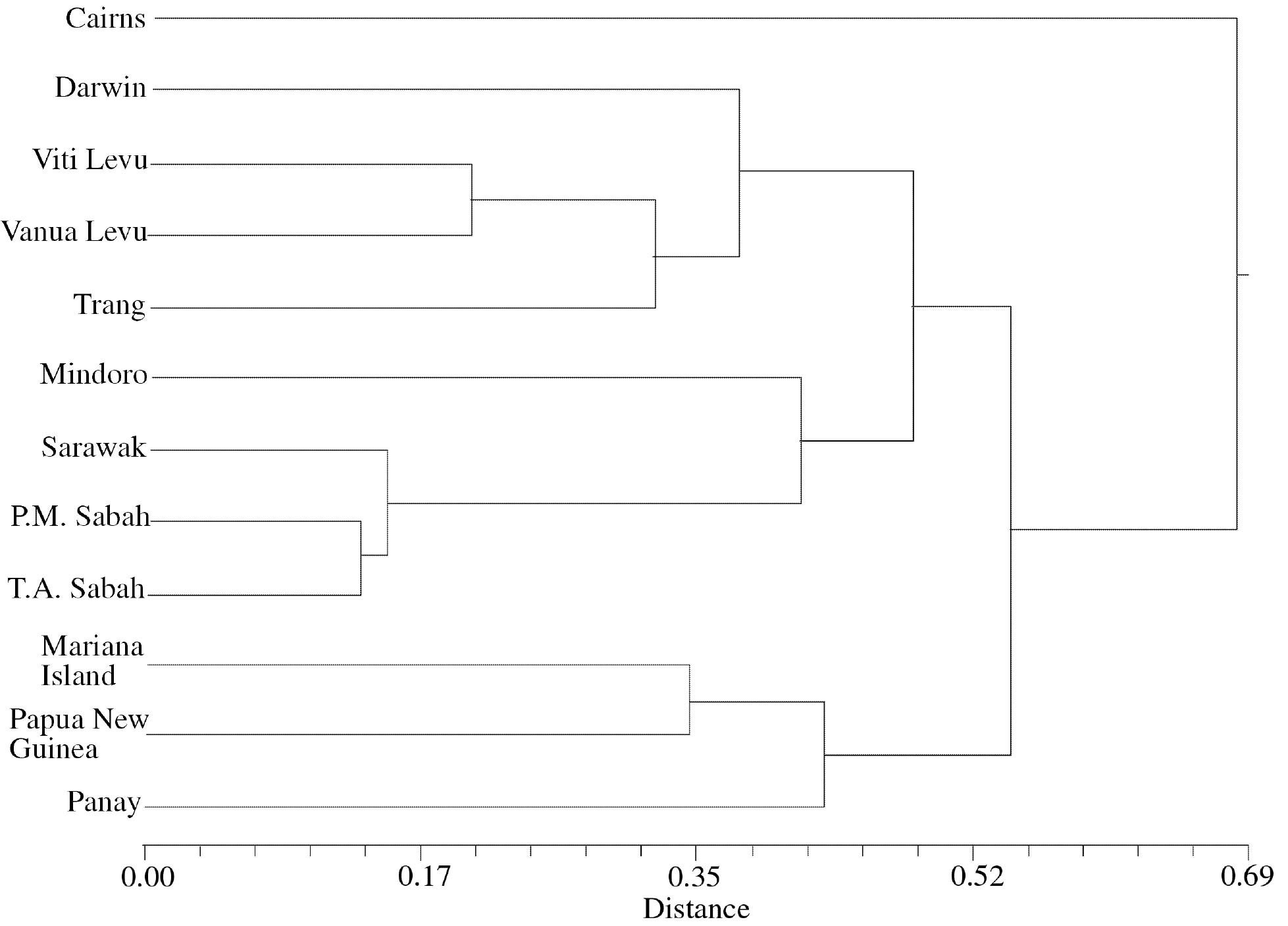
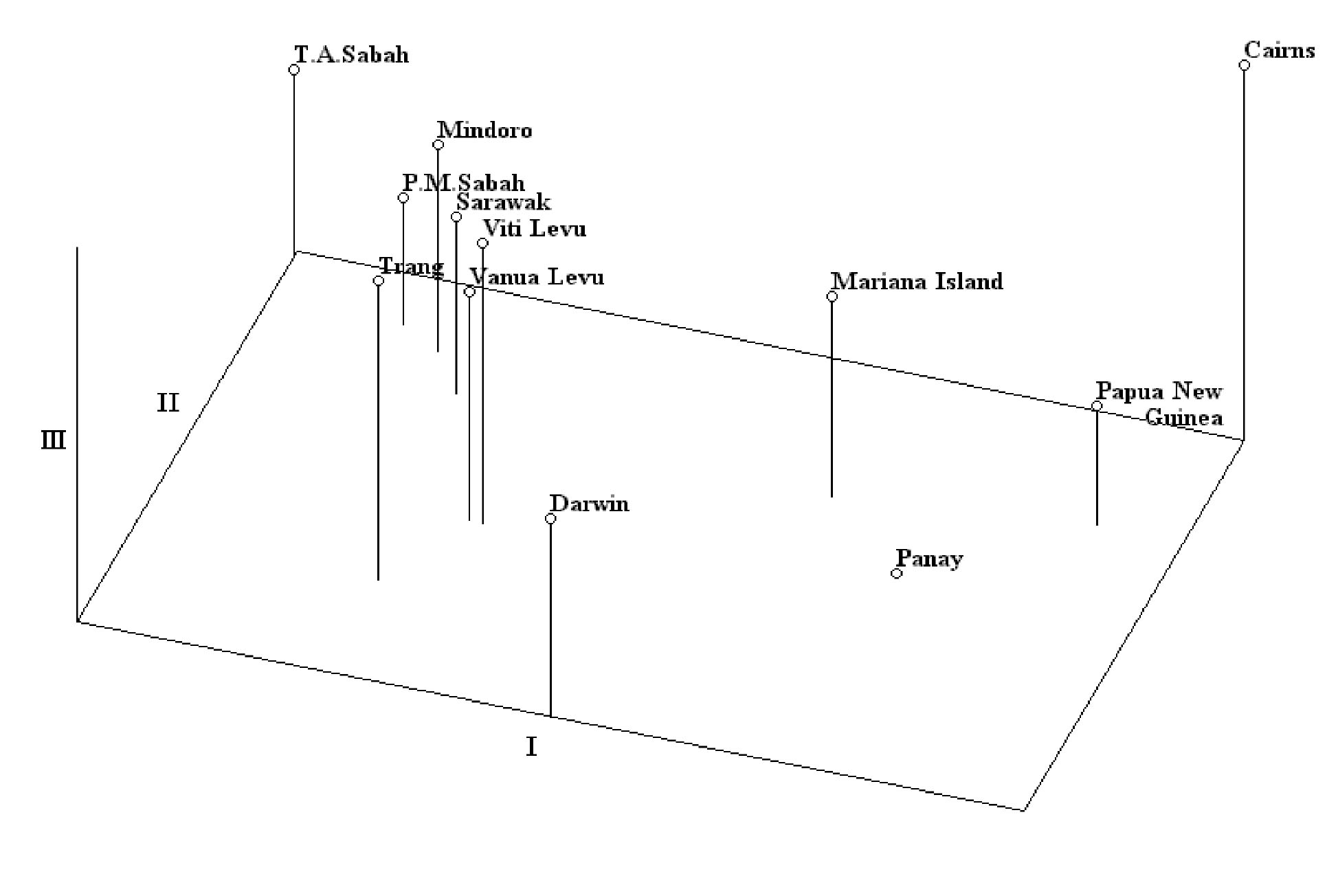
Abstract. One hundred forty-two individual samples belonging to 12 native accessions of Casuarina equisetifolia grown in an international Provenance trial garden in Taiwan were studied for RAPD variation. Twelve primers were used and 89 polymorphic bands were scored. An analysis of molecular variance (AMOVA) revealed that the percentages of variance attributable to variation among and within provenances were 39.28% and 60.72%, respectively. When accessions of north and south hemispheres were treated as two separate groups, the variance components between groups, among provenances within groups, and among individuals within provenances were 2.42%, 37.55%, and 60.03%, respectively. The difference between north and south latitude groups was not significant (p=0.1738). The average Nei's gene diversity for a location of origin was 0.1473. Cluster analysis and principal coordinates analysis revealed two major groupings. One group comprised solely the Cairns accession of Australia and identifies C. equisetifolia var. incana while the other group comprised the remaining accessions of C. equisetifolia var. equisetifolia. The RAPD data support the taxonomic treatment of two varieties in this species. Some regional relationships were observed within C. equisetifolia var. equisetifolia. Study results indicated a large genetic variation among the native accessions of Casuarina equisetifolia. Therefore, the results of provenance trials are expected to be useful in the selection of a suitable provenance for a particular coastal environment.
Keywords: Casuarina equisetifolia; Genetic variation; RAPD; Provenance trial.
Introduction
Casuarina equisetifolia Forst & Forst is the most widely cultivated species within the family Casuarinaceae. It is grown, especially in China and India, for landscaping, pulp, lumber, medicine, tannin, dye, and sand-shifting control in coastal areas (Pan et al., 1996). The species is distributed in Southeast Asia and Australia, which have large native populations, and is introduced into other subtropical and tropical areas (Doran and Hall, 1983; Wilson and Johnson, 1989). The plant was first introduced to Taiwan in 1897, employed as an important pioneer species in coast areas because of its salt and drought tolerance, fast growth, and suitability for sand-shifting control. Other Casuarina species have also been introduced in Taiwan. Introgressive hybridization among species may have occurred (Wang et al., 1984) because of the long cultivation history and the similar flowering periods of different species. Such hybridization can make species identification difficult (Hwang and Hsiao, 1985; Badran et al., 1976).
In 1992, Taiwan Forestry Research Institute participated in an international cooperation program, which conducted a Casuarina equisetifolia provenance trial under the Forestry / Fuelwood Research and Development Project (F/FRED) of the WINROCK International Agriculture Development Research Institute and established a Casuarina equisetifolia provenance trial garden at the Suhu station on the western coast of Taiwan. Several other countries including Indonesia, Sri Lanka, Malaysia, and Thailand also participated in this program. The program was funded by the International Development Foundation in the United States and was overseen by the Commonwealth Scientific and Industrial Research Organization (CSIRO) of Australia. The seed sets were received from the program and were planted according to the standardized experiment design (Kamis, 1992). The purpose of this international project was to evaluate phenotypic responses to various habitats, to select a suitable provenance for a particular coastal environment, and to study the population genetic variation of the species. Twenty-eight provenances were included in this project. Among these 16 seed sets were collected from cultivated areas while 12 were collected from native populations. The latter were studied to evaluate the genetic variation of the native populations of the species.
*Corresponding author. Tel: 886-4-22840417 ext. 315; Fax: 886-4-22874740; E-mail: jyhsiao@dragon.nchu.edu.tw