Bot. Bull. Acad. Sin. (2002) 43: 107-113
Huang and Liu Carbohydrate metabolism in rice callus
Carbohydrate metabolism in rice during callus induction and shoot regeneration induced by osmotic stress
Wen-Lii Huang1 and Li-Fei Liu2,*
1Department of Biotechnology, Fooyin Institute of Technology, Kaohsiung Hsien 831, Taiwan
2Department of Agronomy, National Taiwan University, Taipei 106, Taiwan
(Received August 8, 2001; Accepted November 5, 2001)
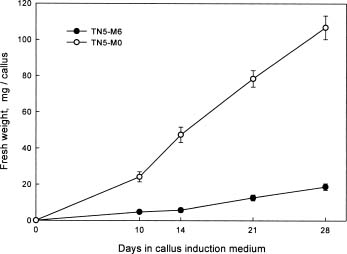
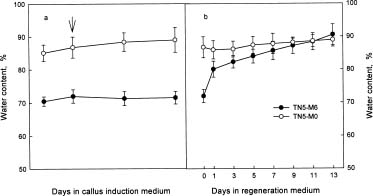
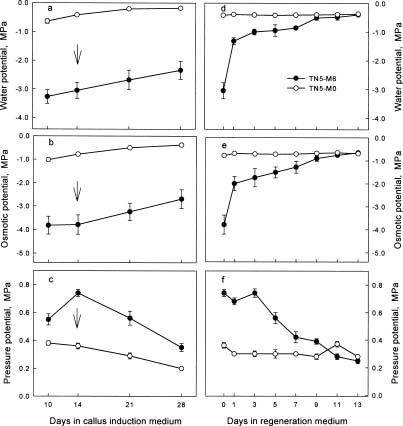
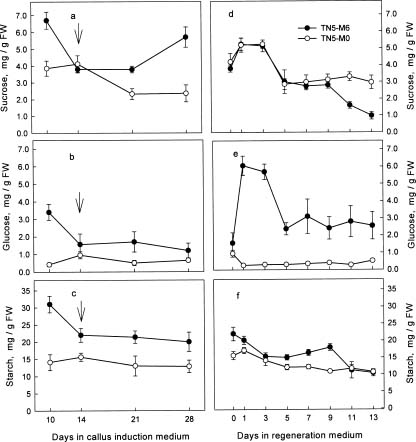
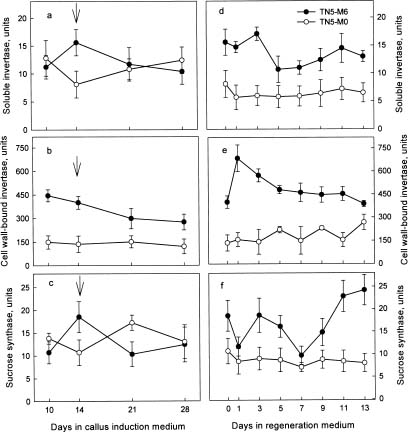
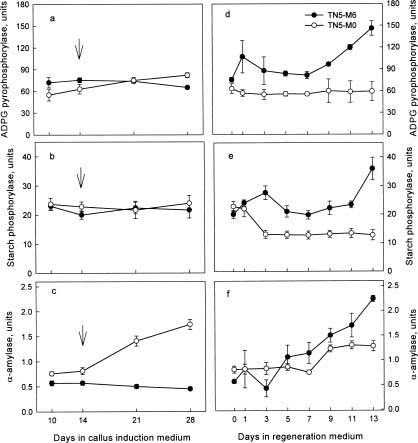
Abstract. We are interested in the cellular physiological events taking place during shoot regeneration in rice (Oryza sativa L. cv. Tainan 5) callus induced by osmotic stress. At first, the sucrose and starch metabolisms in rice callus were studied because carbohydrates are the main energy source in plant tissue culture. The results showed that fresh weight, water content, cellular water, and osmotic potentials all decreased significantly in highly regenerable callus which was induced on MS basal medium supplemented with 10 µM 2,4-D and 0.6 M mannitol (TN5-M6). Besides, the starch and soluble sugar contents in TN5-M6 callus were higher than in un-regenerable callus, induced on the same medium without mannitol. Then, a sudden increase of glucose content was found in TN5-M6 the first day after the callus was transferred to regeneration medium. Simultaneously, the activities of sucrolytic enzymes, sucrose synthase, and acid invertase were higher, and they may have responded to the increase of glucose content. It is suggested that the sudden increase of glucose content may play an important role in shoot regeneration.
Keywords: Carbohydrate metabolism; Oryza sativa; Osmotic stress; Regeneration related factors; Shoot regeneration.
Abbreviations: 2,4-D, 2,4-dichlorophenoxyacetic acid; a-Amy, a-amylase; AGPase, ADP-glucose pyrophosphorylase; Bound-IT, cell wall-bound form invertase; HEPES, N-[2-hydroxyethyl]-piperazine-N'-[2-ethanesulfonic acid]; MS, Murashige and Skoog; RSus, rice sucrose synthase; Sol-IT, soluble form invertase; SPase, starch phosphorylase.
Introduction
Plant cells possess totipotency, i.e., whole plants can be regenerated from single cells by modulating culture conditions (Reinert, 1959). The mechanisms of totipotency, however, are little understood so far, and are mainly discussed in relation to the concentration and ratio of phytohormones (Toonen and De Vries, 1996). It has been reported that osmotic stress affects callus growth, colony formation, shoot regeneration, somatic embryogenesis, and the metabolism of specific compounds (Maretzki et al., 1972; Klenovska, 1973). In previous studies, we discovered that shoot regeneration frequency was dramatically different among rice callus induced from different varieties (Lai and Liu, 1982). Additionally, the shoot regeneration ability of un-regenerable callus could be promoted by osmotic stress treatment (Lai and Liu, 1986; 1988; Liu and Lai, 1991). This provides an alternative concept that the growth and differentiation of cells could be modulated by the cellular physiological water status. We are thus interested in what cellular physiological events occurred during this process.
Carbohydrate supplied to a medium not only acts as a source of carbon and energy, but also as an osmotic agent during organogenesis (Thorpe and Murashige, 1970; Verma and Dougall, 1977). However, very little is known about carbohydrate metabolism in cultured cells. Our preliminary histological study showed that starch granules increased in highly regenerable rice callus. After being transferred to regeneration medium, the callus was able to regenerate shoots in several days, and those starch granules disappeared (Liu and Lee, 1996). The correlation between starch metabolism and shoot formation was reported in tobacco (Thorpe and Murashige, 1968; Thorpe and Meier, 1974; Thorpe et al., 1986), sugarcane (Ho and Vasil, 1983), and Begonia (Mangat et al., 1990). However, there is no further information about carbohydrate metabolism in rice callus. Moreover, no link between osmotic stress, carbohydrate metabolism, and shoot regeneration has been explored.
In this study, callus growth and cellular water status under osmotic stress were measured. Then, the contents of carbohydrates and the activities of enzymes related to sucrose and starch metabolism during callus induction and shoot regeneration were further examined, to clarify the relationship between osmotic stress, carbohydrate metabolism, and shoot regeneration in rice callus.
*Corresponding author. Tel: 886-2-23633502 ext. 101; Fax: 886-2-23633502 ext.123; E-mail: lfliu@ccms.ntu.edu.tw