Bot. Bull. Acad. Sin. (2002) 43: 131-138
Whang et al. A morphometric analysis within the Ixeris chinensis complex
A morphometric analysis of infraspecific taxa within the Ixeris chinensis complex (Asteraceae, Lactuceae)
Sung Soo Whang1, Kyung Choi2,5, Robert S. Hill3, and JaeHong Pak4,*
1Division of Science Education, Chonbuk National University, Chonju 561-756, Republic of Korea
2Department of Biology, Yeungnam University, Keongbuk 712-749, Republic of Korea
3Department of Environmental Biology, University of Adelaide, SA 5005, Australia
4Department of Biology, Kyungpook National University, Taegu 702-701, Republic of Korea
(Received July 17, 2001; Accepted October 3, 2001)
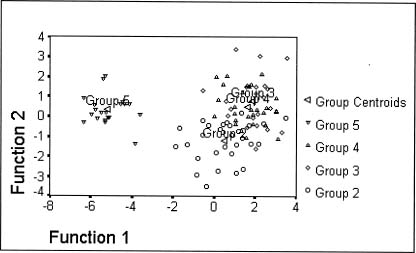
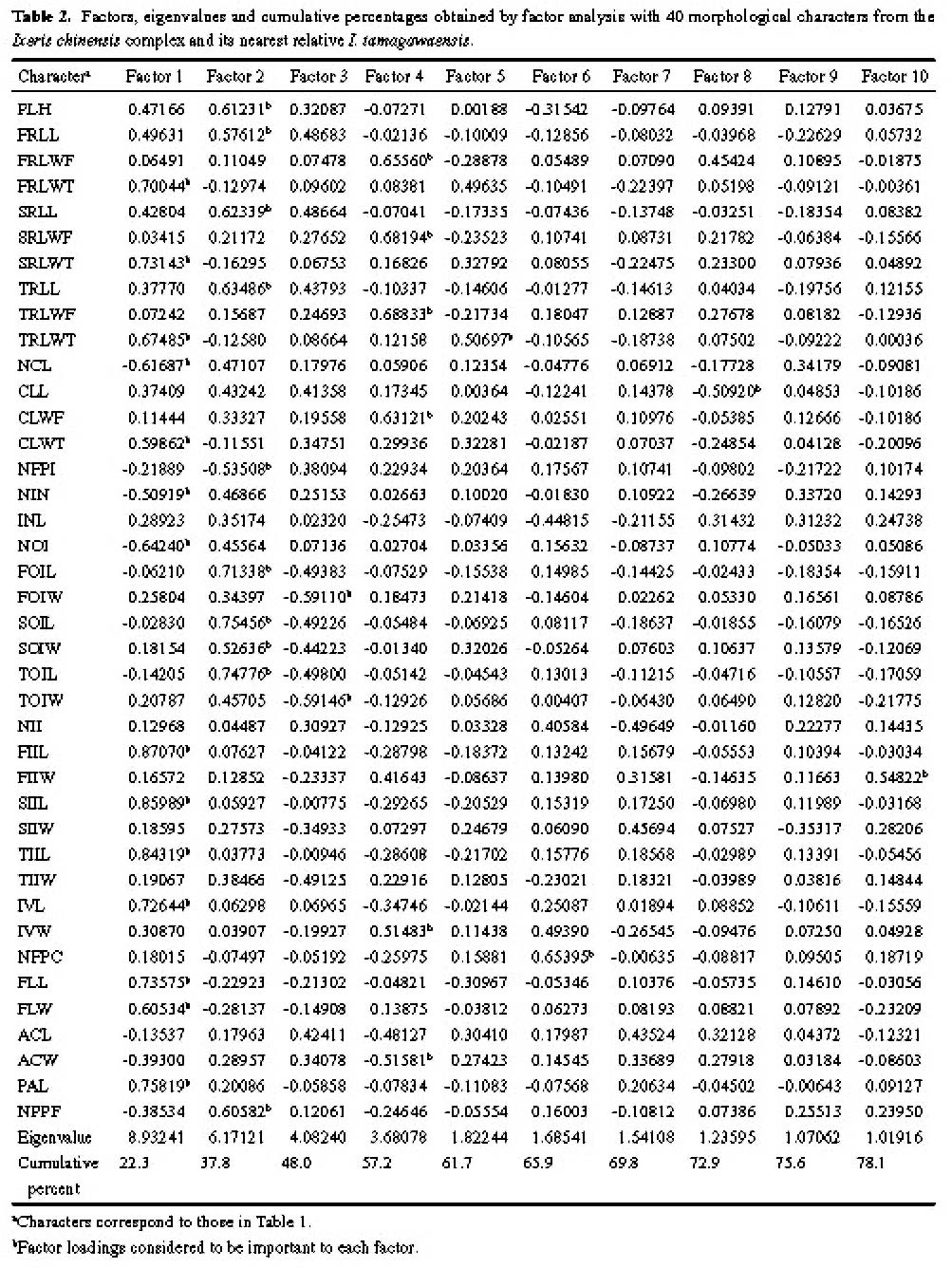
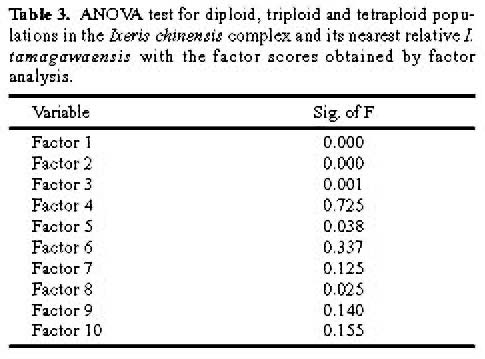
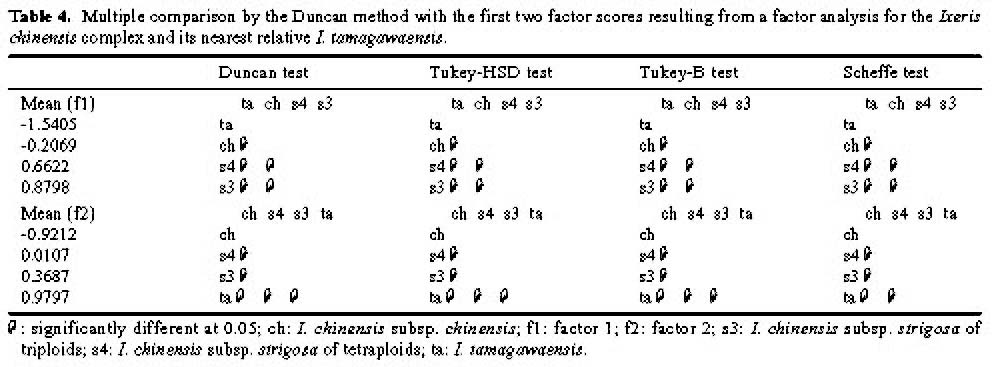
Abstract. Morphometric methods were used with 40 morphological characters from each of 110 individuals collected from 11 populations of the Ixeris chinensis complex to test hypotheses regarding infraspecific taxa. Individuals of the nearest relative I. tamagawaensis were included as an outgroup. To utilize structual associations of the original data and the cumulative values of character variation, a modified data set was generated by matching factor scores with the large original data set. As a result, the original data was reduced to about 10% of its original size. The modified data also emphasized the cumulative values of character variations. Structural associations among characters measured, however, were not significant for most factors extracted even though some obvious trends appeared. Relationships between cumlative values of variation and taxa in the complex are discussed based on the results of the canonical discriminant function analysis, ANOVA, multiple comparison test, and cluster analysis, using the modified data. This study supports Kitamura's (1956) classification of infraspecific taxa in the I. chinensis complex. Given that there is no overlap of cumulative values of variation between the species complex and I. tamagawaensis, there is probably a 75% character overlap within the complex.
Keywords: Ixeris chinensis complex; Infraspecific taxa; Morphometric analysis.
Introduction
The term "species complex" has frequently been applied to flowering plants, but its origin is obscure. It has often been used to describe species aggregations made up of infraspecific taxa such as microspecies, agamospecies, and semispecies (e.g. Fosberg, 1942; Sylvester-Bradley, 1952; Davis and Heywood, 1963; Northington, 1976; Grant, 1981). Several criteria have been used to establish groups of infraspecific taxa, including their morphological discreteness, ecological differences, genetic divergence, and natural reproductive isolation (e.g. Davis and Heywood, 1963; Gilmartin, 1974; Grant 1981; Stuessy, 1990). Morphological discreteness plays a major role in the early stage recognition of infraspecific taxa. A practical difficulty here is that almost all morphological characters and/or character states used for evaluating infraspecific taxa are only slightly differentiated from one another and usually show considerable overlap. Therefore the identification of infraspecific taxa and their hierarchical arrangement within a complex of species usually requires a combination of
several morphological characters, especially if other forms of data are not available.
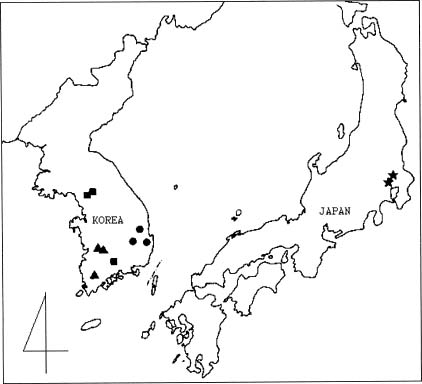
The Ixeris chinensis complex (Asteraceae, Lactuceae) is distributed from Siberia to Japan, including Korea, Taiwan, and China (Pak et al., 1997). This complex has a basic chromosome number of eight (Pak and Kawano, 1990), but also contains diploids (2n = 16), triploids (2n = 24), and tetraploids (2n = 32) (Ishikawa, 1921; Babcock et al., 1937; Chuang et al., 1962; Hsu, 1967, 1970; Peng and Hsu, 1978; Pak and Kawano, 1990) and exhibits sympatry and/or allopatry among the ploidy (e.g. Pak et al., 1995, 1997). The diploid plants observed are sexual while triploids and tetraploids are asexual. Many complicated variations in external morphology have been reported in hybrids between sexual and usually asexual plants, which can revert to sexual reproduction under some conditions (e.g. Harlan and de Wet, 1963; Yahara, 1983, 1990). Within this complex a complicated morphological overlap, without any discontinuities, has led to taxonomic difficulty (Kitamura, 1956; Pak and Kawano, 1992; Kim et al., 1999). Kitamura (1956) classified this complex into the three subspecies Ixeris chinensis (Thunb.) Nakai subsp. chinensis, subsp. strigosa (H. Lév. et Vaniot) Kitamura, and subsp. versicolor (Fischer) Kitamura. Tzvelev (1964) classified it as three different species in a different genus, Ixeridium chinensis (Thunb.) Tzvelev, I. strigosum (H. Lév. et Vaniot) Tzvelev and I. gramineum (Fisch.) Tzvelev. Pak and
5Current address. Division of Life Science & Graduate School of Biotechnology, Korea University, Seoul 136-701, Republic of Korea.
*Corresponding author. Tel: +82-53-950-5352; Fax: +82-53-953-3066; E-mail: jhpak@kyungpook.ac.kr