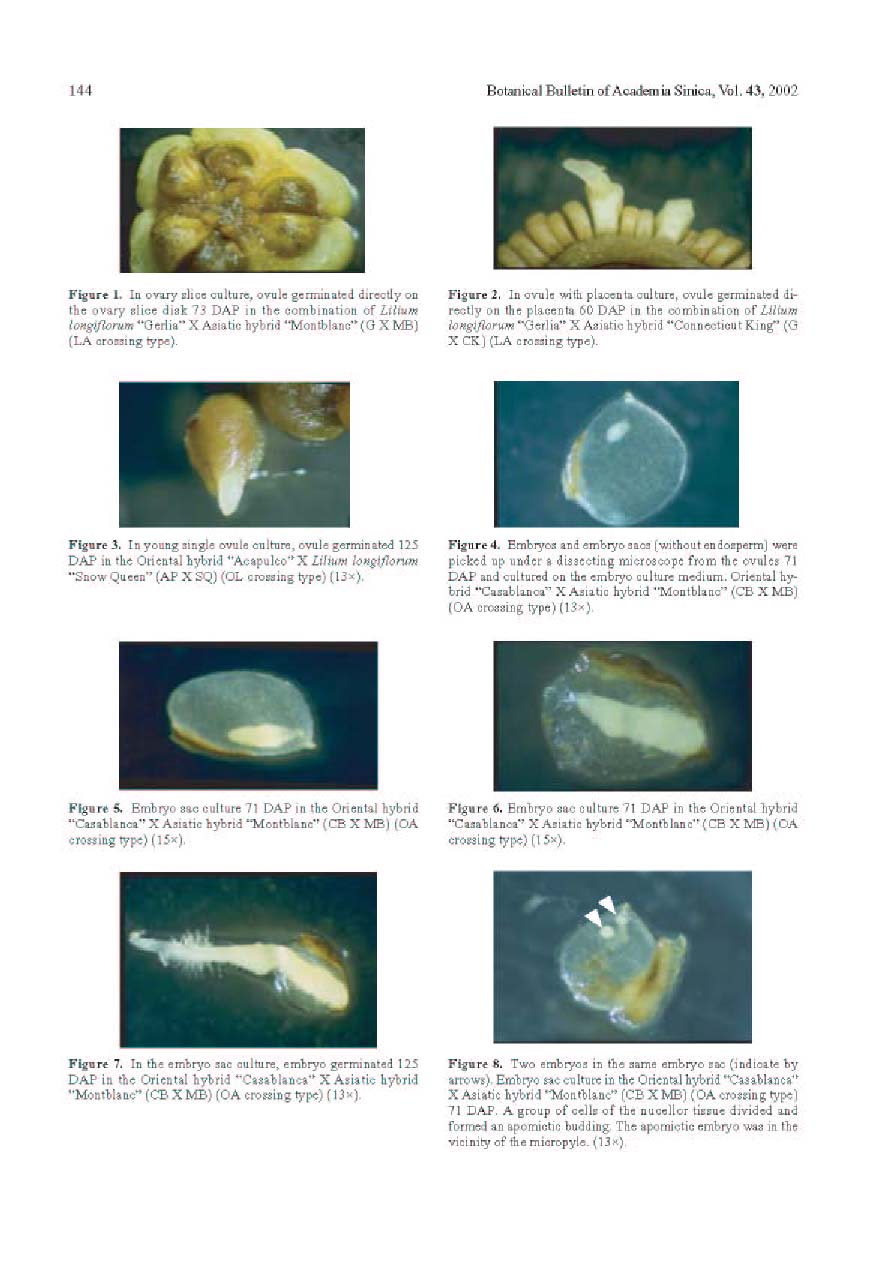
Bot. Bull. Acad. Sin. (2002) 43: 139-146
Chi Various embryo rescue methods in interspecific crosses of lily
The efficiencies of various embryo rescue methods in interspecific crosses of Lilium
Hai-Shan Chi
Department of Horticulture, National Chiayi University, Chiayi 600, Taiwan, Republic of China
(Received July 16, 2001; Accepted January 22, 2002)
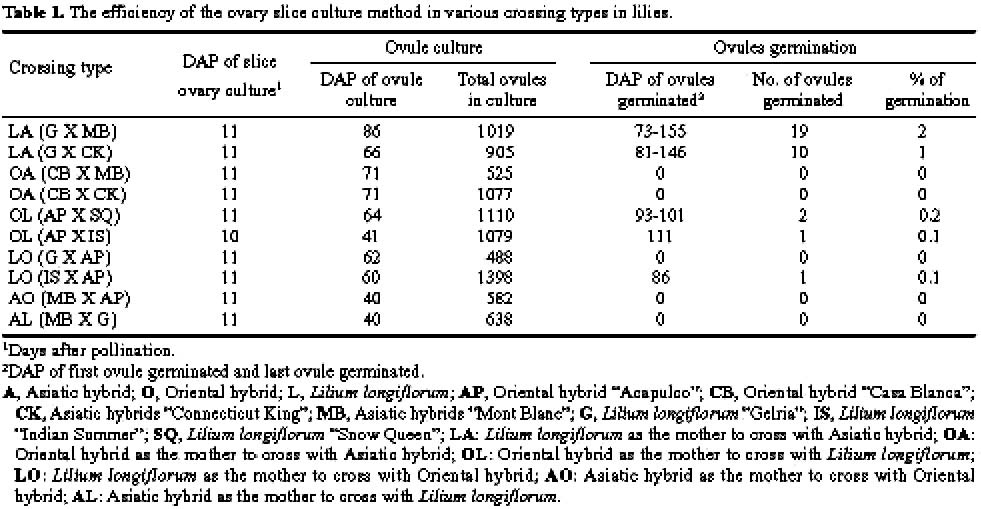
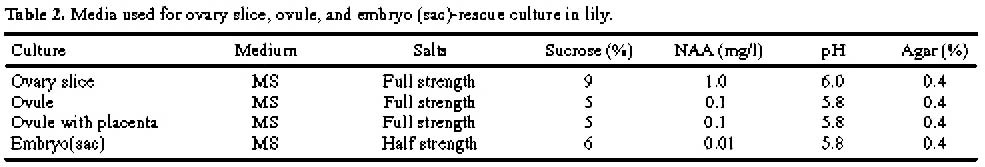
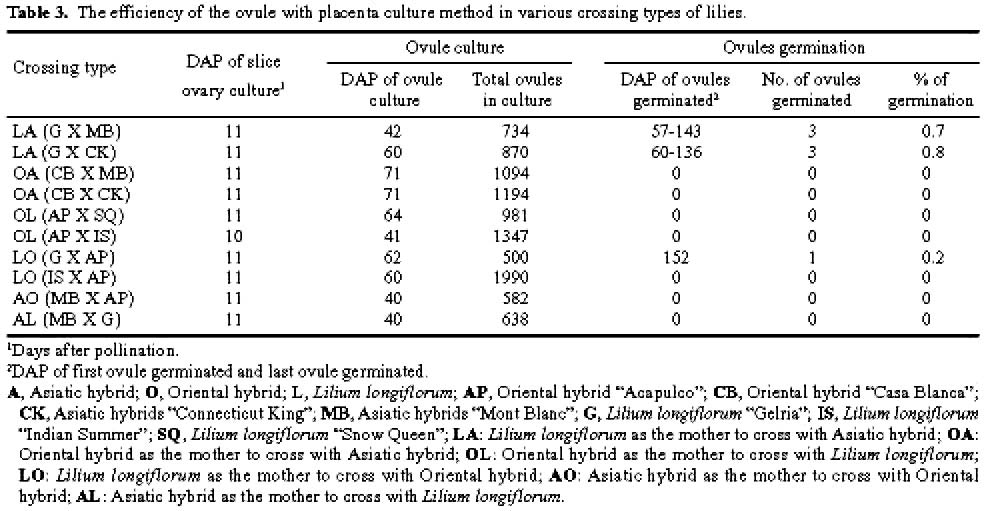
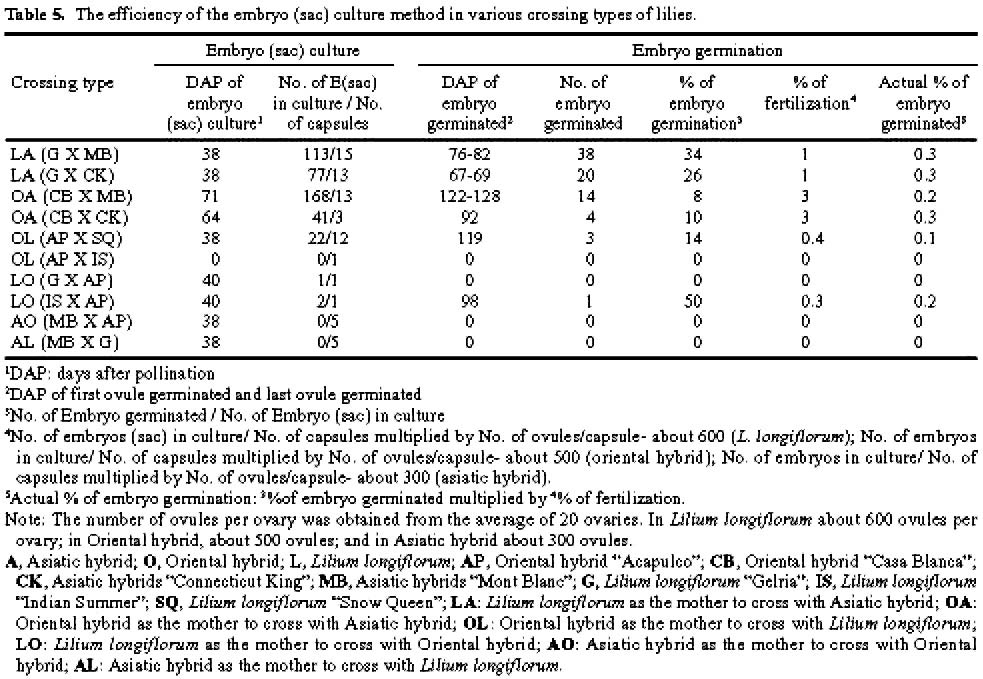
Abstract. The purpose of this research was to compare the efficiencies of various embryo rescue methods in interspecific hybridization of lily. This research included ten combinations of crosses with six crossing types (Lilium longiflorum X Asiatic hybrid, LA; Oriental hybrid X Asiatic hybrid, OA; Oriental hybrid X Lilium longiflorum, OL; Lilium longiflorum X Oriental hybrid, LO; Asiatic hybrid X Oriental hybrid, AO; and Asiatic hybrid X Lilium longiflorum, AL). Four culture methodsovary slice culture (OSC), ovule with placenta culture (OPC), young single ovule culture (YOC), and embryo (sac) rescue method (ESR)were used to culture the fertilized ovules after cut style pollination on plants. The OSC method could be used to obtain hybrid plants in each of the crossing types (0.1-2% success), except for in the OA, AO, and AL crossings. With the OPC method, ovule germinated (0.2-0.8% success) only when Lilium longiflorum "Gerlia" was used as the maternal parent. OSC and OPC required more complicated manipulations. When using the YOC method, ovules were harvested 10 days after the interspecific crosses, and embryo could be germinated from various genotype combinations (0.1-1.1% success). The number of days after pollination (DAP) required for ovule germination in this method (60-125 DAP) was fewer than in the OSC (73-155 DAP) and OPC (57-152 DAP) methods. More plantlets were obtained by the ESR method. The value of the ESR method is that, in the OA crossing, the embryo could only be germinated (0.2-0.3% success) after about 2-3 months of cut style pollination. Apomixes were found in the ovules in some of the cases in this crossing type. No plantlets were obtained in the AL or AO crossings with any of the culture methods tested. Based on the data obtained here, the ESR appears to be the most effective method for obtaining hybrid plants in interspecific hybridization of lily.
Keywords: Apomictic embryo; Cut style pollination; Embryo (sac) culture; In vitro culture; Interspecies crosses; Lilium; Ovary slice culture; Ovule with placenta culture; Young single ovule culture.
Abbreviations: A, Asiatic hybrid; O, Oriental hybrid; L, Lilium longiflorum; AP, Oriental hybrid "Acapulco"; CB, Oriental hybrid "Casa Blanca"; CK, Asiatic hybrids "Connecticut King"; MB, Asiatic hybrids "Mont Blanc"; G, Lilium longiflorum "Gelria"; IS, Lilium longiflorum "Indian Summer"; SQ, Lilium longiflorum "Snow Queen"; DAP, days after pollination; E (sac), embryo and/or embryo sac; CSM, cut style pollination method; OSC, ovary slice culture; OPC, ovule with placenta culture; YOC, young single ovule culture; ESR, embryo (sac) rescue method.
Introduction
In the genus Lilium, interspecific hybridization has been conducted to produce novel hybrids which can combine the Botrytis resistance of Orientals, the virus resistance of Asiatics, and the Fusarium resistance of trumpets with attractive blooms and good growing qualities. Various techniques have been applied successfully to overcome pre- and post-fertilization of incompatibility. A comparison of various pollination methods concluded that pre-fertilization barriers could be overcome by using the cut style technique (Van Tuyl et al., 1982, 1988). This method applies the pollen on the stylar surface after removing the stigma and part of the style, which inhibit pollen tube growth. Once fertilization has occurred, the growth of the hybrid embryos may still be restricted by post-fertilization barriers. These may be partly overcome by using the embryo rescue method in lily as described by several authors (North and Wills, 1969; Asano and Myodo, 1977; Asano, 1980; Van Tuyl et al., 1986).
While post-fertilization barriers may be overcome by placenta culture (Janson, 1993; Niimi et al., 1995) and ovary slices may be cultured successfully starting 40 days (Kanoh et al., 1988) or 5-8 days after pollination (Van Tuyl et al., 1991), the integrated use of cut-style pollination followed by embryo rescue has generated some new interspecific hybrids (Van Tuyl et al., 1991). A drawback associated with the cut-style method, however, is the low seed set (Van Tuyl et al., 1982, 1986), presumably caused by the premature arrival of pollen tubes in the ovary (Janson et al., 1993) and the low rate of pollen tube penetration into the micropyle of ovules instead of sperm cells formation (Chi, 2000). Van Tuyl et al. (1991) suggested that the low number of hybrid plants recovered might be attributable to the relatively long interval between pollination and embryo rescue (Van Tuyl et al., 1991). A simple method for obtaining seedlings from very young embryos was established in L. formosamum by the young single ovule culture (YOC) technique (Niimi et al., 1995). Niimi et al. (1995) found that seedlings could be obtained from