Bot. Bull. Acad. Sin. (2002) 43: 147-154
Liu et al. Species pairs of the Podophyllum group
Molecular evidence for the sister relationship of the eastern Asia-North American intercontinental species pair in the Podophyllum group (Berberidaceae)
Jianquan Liu1,2,*, Zhiduan Chen2, and Anming Lu2
1Northwest Plateau Institute of Biology, the Chinese Academy of Sciences, Qinghai 810001, P.R. China
2Laboratory of Systematic and Evolutionary Botany, Institute of Botany, the Chinese Academy of Sciences, Beijing 100093, P.R. China
(Received December 30, 2000; Accepted November 22, 2001)
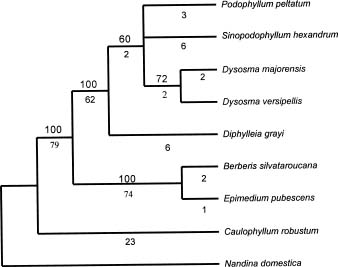
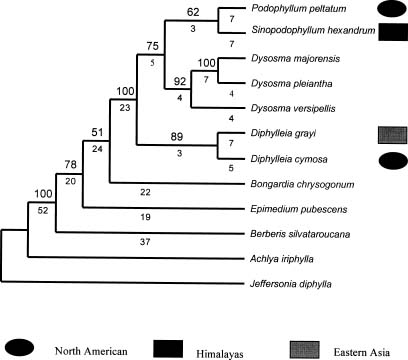
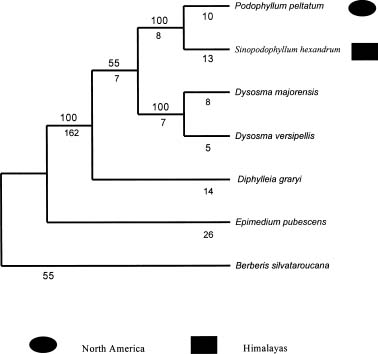
Abstract. The presumed pair relationships of intercontinental vicariad species in the Podophyllum group (Sinopodophyllum hexandrum vs. Podophyllum pelatum and Diphylleia grayi vs. D. cymosa) were recently considered to be paraphyletic. In the present paper, the trnL-F and ITS gene sequences of the representatives were used to examine the sister relationships of these two vicariad species. A heuristic parsimony analysis based on the trnL-F data identified Diphylleia as the basal clade of the other three genera, but provided poor resolution of their interrelationships. High sequence divergence was found in the ITS data. ITS1 region, more variable but parsimony-uninformative, has no phylogenetic value. Sequence divergence of the ITS2 region provided abundant, phylogenetically informative variable characters. Analysis of ITS2 sequences confirmeda sister relationship between the presumable vicariad species, in spite of a low bootstrap support for Sinopodophyllum hexandrum vs. Podophyllum pelatum. The combined ITS2 and trnL-F data enforced a sister relationship between Sinopodophyllum hexandrum and Podophyllum pelatum with an elevated bootstrap support of 100%. Based on molecular phylogeny, the morphological evolution of this group was discussed. The self-pollination might have evolved from cross-fertilization two times in this group. The different pollination and seed dispersal systems of Sinopodophyllum hexandrum and Podophyllum pelatum resulted from their adaptations to different ecological habitats. The divergence time of Sinopodophyllum hexandrum -Podophyllum pelatum is estimated to be 6.52±1.89 myr based on the ITS divergence. The divergence of this species pair predated or co-occurred with the recent uplift of the Himalayas 4-3 myr during the late Miocene and the formation of the alpine habitats. Sinopodophyllum hexandrum developed a host of specialized characters in its subsequent adaptation to the arid alpine surroundings. The present study confirmed the different patterns of species relationship between Asian-North American disjuncts. The isolation of plant elements between North America and eastern Asia must have been a gradual process, resulting in the different phylogenetic patterns and divergence times of the disjuncts.
Keywords: Berberidaceae; Disjunction; Eastern Asia; North American; Sister group Podophyllum group; Species pair.
Abbreviations: myr, million years ago; ITS, internal transcribed spacer of 18S-26S nuclear ribosomal DNA; trnL-F, trnL (UAA) 5´ exon-trnF (GAA) exon region of chloroplast DNA.
Introduction
The disjunct distribution of morphologically similar plants between eastern Asia and North America has been a classical topic since the Linnean era. Many previous studies have contributed to a general documentation of this pattern in various plant groups (Li, 1952, 1972; Wu, 1983; Tiffney, 1985; Hong, 1993; Wen, 1998). This biogeographic pattern is currently recognized for approximately 91 genera (Hong, 1993), and numerous presumable spe
cies pairs have been proposed between the two geographical areas (Li, 1952, 1972). These vicariad species pairs have recently received great attention because phylogenetic analysis, based mainly on molecular data, has shown most of them to be paraphyletic (Wen et al., 1996, 1998; Wen, 1998, 1999; Prince and Parks, 1998) or polyphyletic (Qiu et al., 1995; Soltis and Kuzoff, 1995; Soltis et al., 1995). Furthermore, the disjuncts in the phylogeny also showed various patterns. In Nyssa (Cornaceae), Wen and Stussy (1993) detected two phylogenetic connections between Asia and the Americas, but in Hamamelis (Hamamelidaceae), two Asian species comprised the basal clade, and four North American species formed the other monophyletic clade (Wen and Shi, 1999). Xiang et al. (1998) revealed that four studied genera (Cornus, Boykinia, Tiarella and Trautvetteria) showed this pattern of biogeo
*Corresponding author. The permanent address: Northwest Plateau Institute of Biology, the Chinese Academy of Sciences, Qinghai 810001, P.R. China. Tel: 86-971-6153387; Fax: 86-971-6143282; E-mail: Ljqdxy@public.xn.qh.cn