Bot. Bull. Acad. Sin. (2002) 43: 227-230
Ho et al. Phytophthora insolita on Hainan Island
Phytophthora insolita on Hainan Island
H.H. Ho1,*, H.C. Zeng2, and F.C. Zheng2
1Department of Biology, State University of New York, New Paltz, New York 12561, USA
2Plant Protection Research Institute, Chinese Academy of Tropical Agricultural Sciences, Danzhou City, Hainan 571737, People's Republic of China
(Received July 17, 2001; Accepted September 27, 2001)
Abstract. Phytophthora insolita was isolated from forest soil, streams, and ponds on southern China's Hainan Island. This is the first documented report of its occurrence outside Taiwan, and the significance of the finding is discussed.
Keywords: China; Hainan Province; Phytophthora; P. insolita.
Introduction
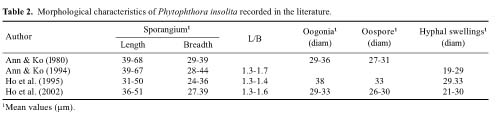
Phytophthora insolita Ann & Ko is an interesting but little known fungus, distinguished from all other species of Phytophthora in the production of abundant oospores in single cultures without the presence of antheridia (Stamps et al., 1990). Although parthenogenic "oospores" have been reported occasionally in P. infestans de Bary and P. fragariae Hickman (Pethybridge and Murphy, 1913; Savage et al., 1968), they are rare and produced only under certain cultural conditions. Phytophthora insolita was first reported from citrus soil in Changhua, Taiwan (Ann and Ko, 1980) and later from diseased poinsettia (Euphorbia pulcherrima Willd.), also in Changhya. Later, the common occurrence of asexual isolates of P. insolita was discovered in soil, ditch water, and diseased plant tissues in southern and central Taiwan (Ann and Ko, 1994). In a recent survey of Phytophthora species on southern China's Hainan Island we have obtained isolates of P. insolita and are reporting them herewith as new findings.
Materials and Methods
In the summers of 1998 and 1999, samples of soil, stream and pond water were collected from natural mountain forests and other localites and brought back to the Plant Pathology Laboratory in Danzhou City for the isolation of Phytophthora species. Samples were placed in 9-inch specimen dishes, with the soil samples flooded with an equal amount of deionized water. Citrus leaves were used as the bait, submerged or partially submerged in water. When lesions started to develop within a few days at room temperature (27-30°C), they were removed, scrubbed slightly with domestic bleaching powder and washed clean under running tap water. Small pieces of plant tissues (ca 2×4 mm) were cut out from the margin the lesions, blotted
dry with paper towels and plated on selective medium in 9-cm petri dishes. The medium was made up by mixing 10 ml Campbell's V-8 juice, 90 ml deionized water, and 0.2 g CaCO3. This was filtered through two layers of cheescloth , diluted with deionized water to 1000 ml, boiled to dissolve 20 gm Bacto agar, and autoclaved at 121°C for 15 min. Prior to pouring the agar medium into sterilized petri dishes, benomyl (150 ppm), hymexazol (50 ppm), rifampicin (100 ppm), nystatin (50 ppm), and ampicillin (100 ppm) were added. The inoculated plates were incubated in dark at 25°C. When mycelial colonies appeared in 2-3 days, they were examined under light microscope to determine their identity as species of Phytophthora based on the colony morphology and the characteristics of the hyphae and branching characteristics (Ho et al., 1995). The culture was then transferred onto 10% V-8 agar plates for further studies. To induce sporangial production, small mycelial agar discs (ca 2×2 mm) were cut from the edge of a growing colony and transferred to sterile distilled water in 6-cm petri dishes, left under regular indoor light and at room temperature.
The pathogenicity of Phytophthora insolita to fruits was tested by inoculating apple, avocado, cucumber, eggplant, mango, green pepper and tomato with the fungus. The fruit was washed clean, surface sterilized with 90% ethanol, and a slit was made with a flamed scalpel. A small mycelial disc from a young colony of isolate Bds 1-18 was inserted into the crevice, which was then sealed with Scotch tape. Alternatively, a small mycelial agar disc (ca 5×5 mm) was placed on an unwounded surface, kept moist with wet cotton, and held in place by Scotch tape. The inoculated fruits were kept in sealed plastic bags and incubated at 28°C for 5 days.
Results
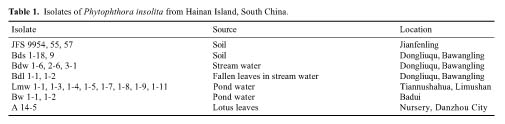
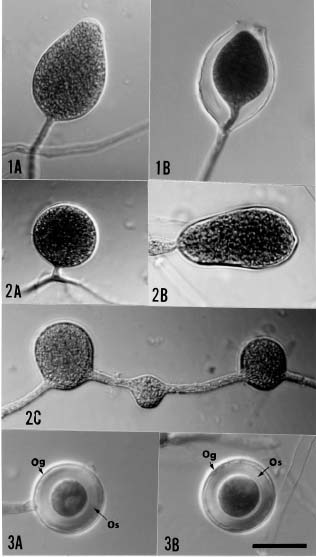
Of the numerous isolates of Phytophthora recovered, those that have been identified as belonging to P. insolita are presented in Table 1. They were all similar in producing appressed and petaloid colonies on V-8 agar plates,
*Corresponding author. Tel: 845-257-3780; Fax: 845-257-3791; E-mail: hoh@newpaltz.edu