Bot. Bull. Acad. Sin. (2002) 43: 251-259
Chen et al. An antimicrobial protein cDNA from mung bean
Cloning and characterization of a cDNA encoding an antimicrobial protein from mung bean seeds
Kuan-Chung Chen1, Ching-Yu Lin1, Mei-Chu Chung2, Cheng-Chun Kuan1, Hsien-Yi Sung1, Samson C.S. Tsou3, C. George Kuo3, and Ching-San Chen1,2,*
1Graduate Institute of Agricultural Chemistry, National Taiwan University, Taipei 106, Taiwan
2Institute of Botany, Academia Sinica, Nankang, Taipei 115, Taiwan
3Asian Vegetable Research and Development Center (AVRDC), P.O. Box 42, Shanhua, Tainan 741, Taiwan
(Received March 20, 2002; Accepted April 16, 2002)
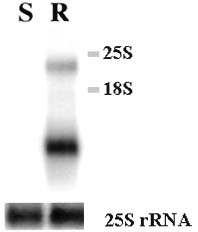
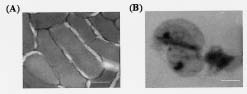
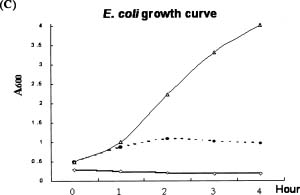
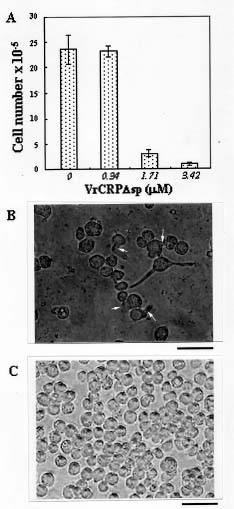
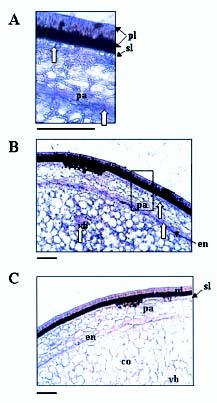
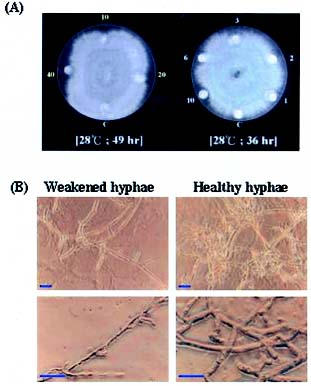
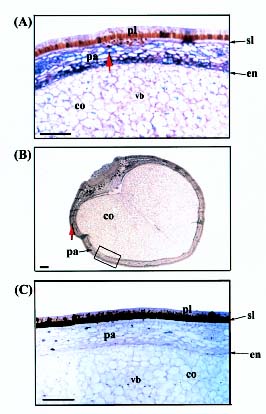
Abstract. A cDNA clone, VrCRP (GenBank accession no. AF326687), encoding an antimicrobial protein was isolated from the differentially expressed cDNAs between a pair of near isogenic lines of mid-matured mung bean seeds. Northern blot analysis showed that VrCRP gene is preferentially expressed in bruchid-resistant developing mung bean seeds. The mature VrCRP is a basic cysteine-rich protein with a predicted molecular mass of 5.9 kDa and a calculated isoelectric point of 8.23. Eight cysteine amino acids present in the primary structure of VrCRP compose four disulfide bridges. Biological analysis showed that VrCRP is toxic to E. coli and completely arrested growth of insect cell (Spodoptera frugiperda, Sf21) at a low concentration of 3.42 µM. VrCRP also inhibited the growth of Rhizoctonia solani. Immunolocalization and in situ hybridization indicated that VrCRP was predominantly present in the parenchyma cells of the seed coat. Based on the biological activities and the protein localization, it is suggested that VrCRP may act as an evolutionary barrier in mung bean seeds that is important in coping with invasions of pathogens and herbivores.
Keywords: Antimicrobial protein; Immunolocalization; In situ hybridization; Mung bean.
Introduction
Plant seeds are usually sown in natural surroundings rich in microorganisms and insects. During evolution, various defense mechanisms developed in plants-such as low water content in the seed and a hard seed coat to provide effective physical defenses against phytopathogens and herbivorous animals. Another array of defense mechanisms includes the production of antimicrobial phytoalexins (Van Etten et al., 1989), pathogenesis-related proteins (Linthorst, 1991; Ponstein et al., 1994), and a large group of cysteine-rich proteins such as lipid transfer protein (Garcia-Olmedo et al., 1995), antimicrobial peptide HvAMP1 (Harrison et al., 1997), Rs-AFP1, Rs-AFP2 (Terras et al., 1992), plant defensins (Broekaert et al., 1995; Terras et al., 1995), hevein (Van Parijs et al., 1991), knottin-type peptides (Cammue et al., 1992; Broekaert et al., 1997), four-cysteine-type peptide (Duvick et al., 1992), 2S albumins (Terras et al., 1992) and thionins (Bohlmann, 1994). The thionins and plant defensins are among the best characterized defensive proteins.
The defensin family is very diverse, and several subfamilies can be recognized on the basis of amino acid sequence homologies (Broekaert et al., 1995), including g-thionin. Plant defensins consist of a triple-stranded an
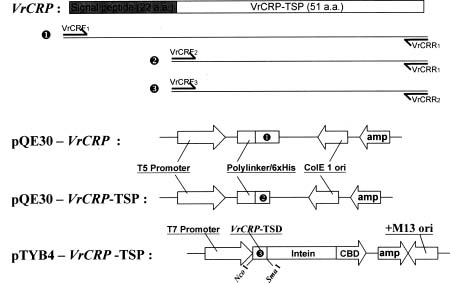
tiparallel b-sheet and a single a-helix lying in parallel with the b-sheet. The Cys-X-X-X-Cys segment of the a-helix is connected by two disulfide bridges to the Cys-X-Cys segment in the third b-strand, forming a cystine-stabilized a-helix motif (Kobayashi et al., 1991). We have isolated a novel cDNA from Vigna radiata, termed VrCRP (Cysteine-rich Protein of Vigna radiata), which encodes a cysteine-rich protein constituting a new member of the plant defensin family. This paper describes the cloning, biological activities, and localization in the seed of this mung bean defensin VrCRP.
Materials and Methods
Plant Varieties
Vigna radiata L. VC1973 and VC6089A were bred in the Asian Vegetable Research and Development Center (AVRDC). Seeds of the former are susceptible to bruchid Callosobruchus chinensis, whereas the latter have been shown to resist bruchid attack. VC6089A (BC6S2) population was bred by crossing a bruchid-resistant wild mung bean, TC1966 (as donor parent) with a breeding line VC1973A (as recurrent parent).
RNA Preparation
Total RNA was extracted from the 15 DAF (days after flowering) developing seeds of VC1973A and VC6089A by hot phenol method (Verwocrd et al., 1989).
*Corresponding author. Tel: 886-2- 2789-9591; Fax: 886-2-2785-8936; E-mail: chingsan@gate.sinica.edu.tw